



Коллектив авторов
Эволюционная и сравнительная психология в России. Теория и практика исследований
Простые системы в психологических исследованиях
Т. А. Палихова, Г. Г. Аракелов
Введение
Российская психология резко тормозит в своем развитии по сравнению с впечатляющими открытиями мировой нейронауки из-за некоторых специфических особенностей своей методологии и особенностей методических подходов к научному исследованию. Опираясь в основном на исследование человека, его психики и поведения, российские психологи упускают уникальную возможность проникнуть в суть, в природу мозговых механизмов обеспечения психики. Основную трудность в интерпретации результатов составляют нечеткость, неконкретность основных терминов. И основная задача ней-ронаук – вдохнуть естественно-научное содержание в такие понятия, как психика, память и т. д. В нейронауках уже стали хрестоматийными результаты, показавшие, что психическое явление появляется при активации определенных нейроэндокринных, распределенных в пространстве и времени нейронных систем. Дальнейшие исследования направлены на уточнение организации конкретных механизмов распределенных систем. Однако исследования на мозге высших животных и человека пока затруднены из-за сложной организации мозга этих существ. Поэтому внимание нейробиологов направлено на простые системы. Простые системы – нервные систем этих животных – относительно просто организованы, но имеют адаптивное поведение.
И. П. Павлов известен всему научному миру благодаря открытию и исследованию им условных рефлексов. Но мало кто знает, что его первая печатная работа называлась «Как улитка раскрывает свои створки» (1885). Она была подготовлена, когда Павлов работал в Физиологическом институте в Бреславле, в лаборатории Р. Гейденгай-на, и была посвящена организации нервного контроля мышечными сокращениями, составляющими поведенческую реакцию моллюска
(Павлов, 1885). Психологи любят ссылаться на системную концепцию П. К. Анохина и мало кто знает, что системный подход у него сформировался как результат попыток изучать генетические основы психики. Он соавтор статьи о роли РНКазы в поведении и обучении (Анохин, Аракелов, Соколов, 1972). Это исследование выполнено на нейронах виноградной улитки. Нейробиологи хорошо знакомы с виноградной улиткой (Helix pomatia L. и Helix lucorum L.) как с популярным объектом нейробиологических исследований. Психологи же знают об этом моллюске (кроме дачных и ресторанных впечатлений) гораздо меньше. Постараемся заполнить этот пробел.
Объект исследования, методы
На кафедре психофизиологии психологического факультета МГУ основной исследовательской стратегией является предложенный
Е. Н. Соколовым подход: Человек – Нейрон – Модель (2003). Исследование нейронов виноградной улитки является необходмой составляющей этого подхода (рисунок 1).

Рис. 1. Безымянный рисунок, взят из одной из интернет-сетей
Во всех учебниках по нейрофизиологии и физиологии ВНД есть главы, посвященные моллюскам. Кроме аксона кальмара как базового объекта клеточной электрофизиологии, обязательно ссылаются на работы Э. Кэндела, лауреата Нобелевской премии (2000) за изучение механизмов синаптической пластичности (Кэндел, 1980). Ссылки на его работы, выполненные на идентфицированных нейронах морского зайца (Aplysia), обязательно присутствуют во всех списках литературы учебников по нейробиологии. Но американская нейронаука родилась в Европе. В том числе Кэндел работал в лаборатории
Ладислава Тока в Париже, где и познакомился с аплизией. К сожалению, в своих публикациях Кэндел на этот факт не ссылается (рисунок 2). Сегодня он увлечен механизмами высших психических функций и искусства (Кэндел, 2016).
Рис. 2. Известная карикатура на Ладислава Тока и Эрика Кэндела
В нашей стране и у соседей исследования на виноградных улитках проводились и проводятся в нескольких институтах и лабораториях: в Киеве, в Институте биофизики под руководством П. Г. Костюка, известного своими электрофизиологическими исследованиями ионных каналов нейрональных мембран; в Казани, в лаборатории Х. Л. Гайнутдинова; и, конечно, в Москве, в институте ВНД и нейрофизиологии РАН, в Институте биологии развития им. Н. К. Кольцова, в Институте психологии РАН и в НИЦ РАН «Курчатовский институт», в отделении нейрофизиологии и когнитивных наук, возглавляемом хорошо известным сегодня нейробиологической аудитории К. В. Анохиным (внуком Петра Кузмича). Многие сотрудники этих лабораторий – выпускники биологического и психологического факультетов Московского университета. Есть и выпускники технических вузов, Физико-технического института и МИФИ, так как интерес биофизиков к вопросам устройства и функционирования мозга растет и будет расти, особенно в связи с нейрокибернетической проблематикой (Соколов, Шмелев, 1985). Выпускники кафедры психофизиологии МГУ работают в институтах других стран. Галина Парамей и сегодня успешно работает в области сенсорной психофизиологии в Hope University в Ливерпуле, а выпускница МИФИ и кафедры Анна Шевякова, живет и работает в Канаде. До сих пор хорошие и продуктивные отношения у нас складывались с литовскими коллегами (Г. Г. Вайткявичюс). Даже на Кубе работают выпускники и аспиранты кафедры. Нельзя не вспомнить Вячеслава Цыганова, выпускника кафедры, выполнившего дипломную работу на нейронах улитки и затем работавшего с В. Е. Дъяконовой и Д. А. Сахаровым в Институте биологии развития на биостанстции под Пущино-на-Оке.
Результаты исследования
Одним из методологических достоинств ««простых нервных систем» является наличие идентифицированных нейронов. Идентифицированные нейроны позволяют исследовать принципиальные вопросы функционирования нервных систем, в том числе принципы организации нейронных сетей, без понимания которых построение «искусственного интеллекта» невозможно. Среди «простых нервных систем» (Балабан, Захаров, 2016) высшие места занимают моллюски и виноградные улитки в том числе. Они привлекают исследователей наличием в их ганглиях индивидуально идентифицируемых нейронов. Это значит, что среди многих разных нервных клеток есть нейроны, которым можно присвоить имя благодаря повторяющимся морфологическим, электрофизиологическим и функциональным показателям (Иерусалимский, Захаров, Палихова и др., 1992). Это уникальное свойство, так как позволяет проводить эксперименты на одних и тех же нервных клетках среди многих разных и не только по типу, но и индивидуально. На данных, полученных на идентифицированных нейронах, построена, например, концепция «командного нейрона», которая и сегодня продолжает обсуждаться. Детальное изучение механизмов организации двигательных реакций позволило показать движения, возникающие в ответ на генерацию одного потенциала действия в командном нейроне (Аракелов, Палихова, 1985).
Идентификация синаптически связанных нейронов позволяет в деталях исследовать синаптические механизмы памяти и научения, которые принято связывать с пластичностью нервной системы. Нобелевская работа Кэндела хорошее тому свидетельство (2000). Е. Н. Соколов предложил для связей между такими нейронами термин «идентифицированные синапсы». Идентифицируемые синапсы изучаются и у виноградной улитки. Так идентифицированы механо-сенсорные нейроны, моносинаптически связанные с командными нейронами париетальных ганглиев. (Аракелов, Маракуева, Палихо-ва, 1989; Malyshev, Balaban, 2002). Их изучение позволило доказать, что синаптическая пластичность участвует не только в процессах памяти, но и сенсорных процессах. Показана, например, динамика амплитуды элементарных возбуждающих постсинаптических потенциалов (ВПСП) во время ответа командного нейрона на меха-носенсорный стимул суммарным ВПСП. Предполагаемое различие между сенсорными нейронами с разной локализацией рецептивных полей открывает перспективу сравнить пластические свойства синапсов с различной эргичностью (холинергичесие и глутаматер-гические). В педальных ганглиях виноградной улитки идентифицированы также нейроны, выполняющие модуляторную функцию с серотонином в качестве медиатора.
Проблема памяти и научения является базовой для исследования идентифицированных нейронов улитки на кафедре психофизиологии. Предложена гипотеза, объясняющая селективность изменения синаптических связей в процессе формирования условного рефлекса. Она получила название «гипотезы пресинаптического ожидания ответа» и заключается в следующем. Повышение уровня кальция в результате спайковой активности пресинаптического нейрона – факт, обязательно представляемый во всех учебниках. Наличие пресинап-тических рецепторов к обратным посредникам, экспрессирующимся при успешной активации постсинаптического нейрона, – тоже. Кальций-зависимое изменение чувствительности рецепторов представляется и обсуждается во многих современных статьях и на конференциях по нейронаукам. Таким образом, повышение чувствительности пресинаптических рецепторов происходит только в предварительно активном нейроне, повышая вероятность селективного влияния обратных посредников только на эти терминали. Доказательство гипотезы – вопрос экспериментальной проверки селективности образования новых связей при обучении. Перспективы – разобраться в механизмах ассоциативного научения, без которых обсуждение высших когнитивных процессов является бессмысленным.
Одним из коррелятов функционального состояния человека являются ритмы мозга. На нейронном уровне не обходится без обсуждения пейсмейкерной активности. Наши исследования показали, что одним из механизмов формирования эндогенных ритмов являются не только свойства каналов нейрональных мембран (рисунок 3), но и такой механизм как «циркуляция возбуждения в нейроне» (см.: Соколов, Шмелев, 1985).
Заключение
В коротком докладе мы попробовали представить данные, полученные на кафедре психофизиологии МГУ на одном из объектов психофизиологических исследований – виноградных улитках, и перечислить вопросы, на которые это животное с «простой нервной системой» может помочь ответить. Очень хотелось бы, чтобы предложенный Е. Н. Соколовым психофизиологический подход «Человек – Нейрон – Модель» развивался и не был заменен «классическими» психофизиологическими исследованиями, в которых ставится вопрос о корреляциях, а не о механизмах психических и психологических процессов.
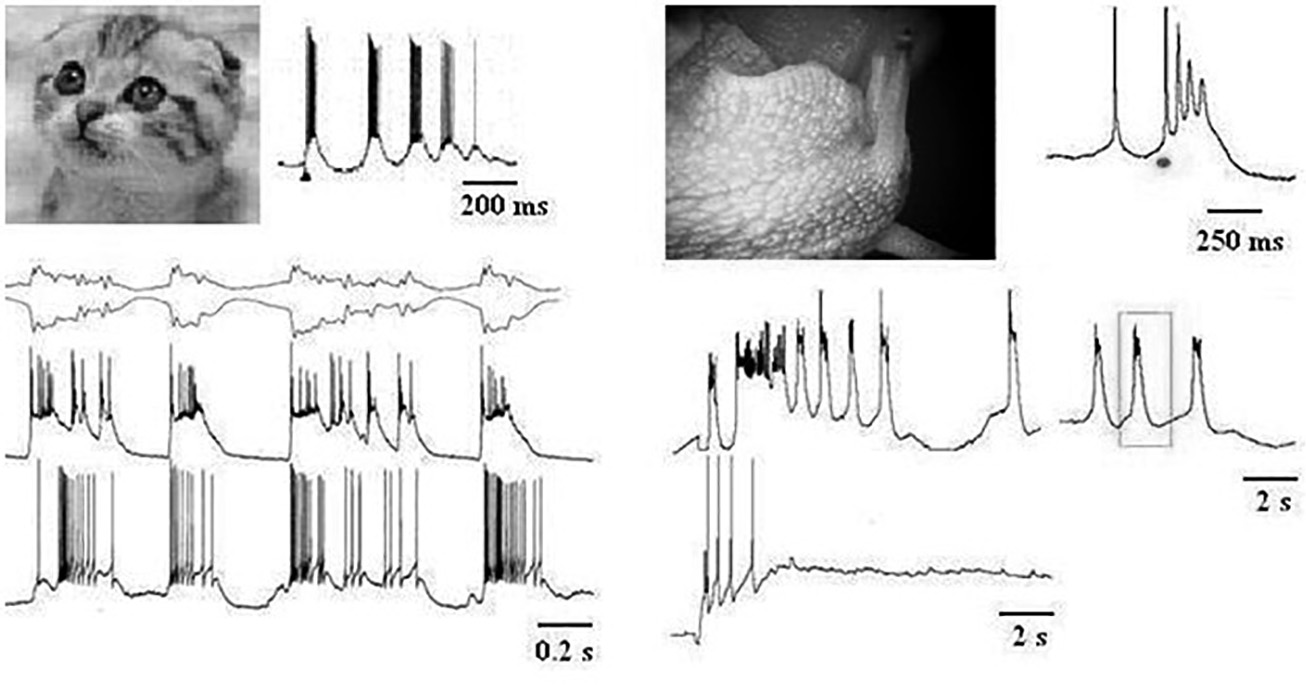
Рис. 3. Внутриклеточно зарегистрированные ритмы в коре кошки и в нейроне улитки У котенка активность нейронов коры и таламуса регистрировалась параллельно с электроэнцефалограммой при частоте пачек и волн ЭЭГ в альфа-диапазоне. В нейроне улитки ритмическая пачечная активность вызвана повышенным содержанием ионов кальция в физиологическом растворе на полуинтактном препарате
Надеемся, что данные, полученные на легочном моллюске виноградной улитки психофизиологическими методами, позволят ответить на вопросы, которые ставит психология перед эволюционной и сравнительной психологией и нейронаукой (Коштоянц, 1957; Сахаров, 1992). И главный среди них: как связаны мозг и психика, в частности в высшем своем проявлении – сознании (Соколов, 2010; Анохин, 2016)?
Литература
Анохин К. В. Коннектом и когнитом: заполнение разрыва между мозгом и разумом. 2016. URL: http://vk.com/kognitivnaja_nauka.
Анохин П. К., Аракелов Г. Г., Соколов Е. Н. Электрогенез в гигантских нейронах моллюска после действия на них рибонуклазы // Нейрофизиология, 1972. Т. 4. № 4. С. 423–4.
Аракелов Г. Г., Маракуева И. В., Палихова Т. А. Моносинаптическая связь: идентифицируемые синапсы в ЦНС виноградной улитки // Жур. ВНД. 1989. Т. 39, № 4. С. 737–745.
Аракелов Г. Г., Палихова Т. А. Центральные механизмы организации движений // Е. Н. Соколов, Л. А. Шмелев (ред.). Вопросы кибернетики. М.: Наука, 1985. С. 84–101.
Балабан П. М., Захаров И. С. (орг.). Простые нервные системы 2016. XI региональная конференция Международного общества нейробиологии беспозвоночных. Москва – Звенигород, 15–19 мая 2016.
Иерусалимский В. Н., Захаров И. С., Палихова Т. А., Балабан П. М. Нервная система и картирование нейронов брюхоногого моллюска Helix lucorum L // Жур. ВНД. 1992. Т. 42. № 6. С. 1075–1090.
Коштоянц Х. С. Основы сравнительной физиологии. Т. 2. Сравнительная физиология нервной системы. М.: Изд-во АН СССР, 1957,
Кэндел Э. Клеточные основы поведения. М.: Мир, 1980.
Кэндел Э. Век самопознания: поиски бессознательного в искусстве и науке с начала ХХ века и до наших дней. М.: ACT – Corpus, 2016.
Павлов И. П. Как беззубка раскрывает свои створки. Опыты и вопросы к общей мышечной и нервной физиологии // Pfluger’s archive. Bd. XXXVII. 1885. S. 36–54. Перевод: И. П. Павлов. Избранные труды / Под. ред. Э. А. Асратяна. М.: Акад. пед. наук РСФСР, 1951. С. 467–493.
Сахаров Д. А. Долгий путь улитки // Жур. ВНД. 1992. Т. 42. № 6. С. 1059–1063.
Соколов Е. Н. Восприятие и условный рефлекс: новый взгляд. М.: УМК «Психология» – Московский психолого-социальный институт, 2003.
Соколов Е. Н. Очерки по психофизиологии сознания. М.: МГУ, 2010.
Соколов Е. Н., Шмелев Л. А. (ред.). Вопросы кибернетики: нейробиологический анализ механизмов поведения. М.: Наука, 1985.
Человек – Нейрон – Модель. Международная научная конференция памяти Е. Н. Соколова и Ч. А. Измайлова. Сборник материалов. 2016. М.: МГУ им. М. В. Ломоносова.
Malyshev A. Y., Balaban P. M. Identification of mechanoafferent neurons in terrestrial snail Helix lucorum L.: characterization of its response properties and synaptic connections // Neurophysiol. 2002. V. 87 (5). P. 2364–2371.
Понятие «инстинкт» и его использование в психологии животных и человека
Г. Г. Филиппова
Проблема инстинкта традиционно считается одной из самых сложных и дискуссионных в зоопсихологии и психологии человека, и тем более она сложна в плане сравнения психики животных и человека (Фабри, 1993; Филиппова, 2012). В современной зоопсихологии и биологии поведения практически исчезло понимание психического компонента инстинкта, реально осталось только инстинктивное поведение. В психологии человека понятие «инстинкт» используется либо для описания неосознаваемого, импульсивного, неконтролируемого поведения, либо как определение «животного», врожденного, а иногда интуитивного чувства (Психологическая энциклопедия, 2003). Недостаточная определенность одного из базовых понятий науки сама по себе требует внимания и анализа, но кроме этого есть насущная необходимость внести ясность в проблему инстинкта в связи с тем, что это понятие активно применяется в психологии человека для объяснения таких феноменов, которые на самом деле не всегда имеют отношение к инстинкту или не имеют такого отношения вовсе.
В качестве примера можно привести представления о репродуктивной функции человека, где активно используются словосочетания «инстинкт размножения или продолжения рода», «материнский инстинкт» и даже «отцовский инстинкт». Со ссылкой на свойство инстинктивного поведения спонтанно проявляться в ответ на ключевой стимул постулируется «врожденная потребность» к материнству и ее проявление при рождении ребенка. При этом в первую очередь имеется в виду чувство матери к ребенку и стремление о нем заботиться. Отсутствие такового и тем более девиантное материнское поведение трактуется как нарушение врожденного влечения (Филиппова, 1998, 2015). При лечении бесплодия в медицине априори считается, что женщина имеет такое врожденное влечение к рождению детей и заботе о них, и игнорируется возможность отсутствия реального влечения к ребенку, что может стать основой внутриличностного конфликта и появления психологической защиты в форме соматизации (нарушений репродуктивного здоровья) (Филиппова, 2015). И тем более это может стать причиной девиантного материнского поведения.
Такое положение требует тщательного анализа самого понятия «инстинкт» и возможности его использования в психологии человека и зоопсихологии.
Источником представлений об инстинкте считается различение «низшей» (у животных) и «высшей» (у человека) формы души в античной философии. Основываясь на учении Платона, Аристотель приписывал человеку «разумную душу», а животным – «чувственную». Чувственная душа животных побуждает их к самосохранению и продолжению рода на основании желаний и влечений, ощущений удовольствия или боли. Само поведение животных Аристотель разделял на врожденное и приобретенное (Философская энциклопедия, 1962; Фабри, 1993; Филиппова, 2012). Таким образом, уже в учении Аристотеля содержится указание на два компонента, которые впоследствии стали основными в анализе инстинктивного поведения: чувственный компонент, который побуждает поведение, и само поведение.
Первое определение инстинкта встречается в философии стоиков и приписывается Хризиппу: инстинкт – это прирожденное влечение, направляющее поведение животных на приятное, полезное и уводящее его от вредного и опасного, которое ощущается как неприятное. Уже в учении стоиков достаточно четко разделяются два базовых компонента инстинкта: прирожденное влечение, которое переживается как чувство и направляет поведение, и инстинктивные действия, которые также считались врожденными, совершались животными без осознавания их смысла и были одинаковыми у всех особей одного вида. В этих определениях заложено понимание психического компонента инстинкта: животное не осознает пользы (биологической) своего поведения, но руководствуется влечением, т. е. переживанием удовольствия и неудовольствия, которое и «ведет» его по правильному пути. Само влечение (т. е. способность испытывать удовольствие и страдание при разных воздействиях и в результате своих действий) является врожденным. Можно сказать, что в этом отношении стоики оказываются более «психологичными», чем современные исследователи инстинкта, практически отрицающие возможность проникновения в субъективный мир животных и сосредотачивающиеся на изучении только поведенческого компонента инстинкта или его физиологических механизмах (Филиппова, 2012).
В дальнейшем, вплоть до начала XIX в. в трудах французских философов-натуралистов (Ж.-О. Ламетри, Э. Б. Кондильяк, Ш. Ж. Леруа, Ж. Л. Л. Бюффон и др.) разрабатывалась проблема различий врожденного и приобретенного поведения у животных и наличия у них способностей, сходных с разумом человека (Фабри, 1993; Филиппова, 2012). Основной задачей стало понимание соотношения инстинкта, научения («привычек») и разумного поведения. В этот период развивался объективный подход к исследованию поведения животных и критиковалась интерпретация их состояний по аналогии с человеческими чувствами. Основным стало понимание приспособительной функции поведения и его изменения в эволюции видов. Такой подход надолго заложил традицию объективного изучения психики и поведения животных и отодвинул в тень компонент влечения в структуре инстинкта.
К чувственному компоненту инстинкта в начале XIX в. вновь обратился Ж. Б. Ламарк. Он определял инстинкты животных как наклонность, вызываемую ощущениями на основе возникших в силу их потребностей и понуждающих к выполнению действий без всякого участия мысли и воли (Философская энциклопедия, 1962; Фабри, 1993). Мысль и воля – это привилегия человека, но чувства и влечения – это компонент инстинктивного поведения животных. Однако основным все же оставалось изучение инстинктивного поведения.
Следующий этап развития учения об инстинктах связан с учением Ч. Дарвина, который, с одной стороны, сравнивал инстинкты животных и человека именно на основании сходства их чувственного, эмоционального компонента, а с другой – обосновал изучение инстинктов как компонента эволюционного процесса (Дарвин, 1953). С этого времени в изучении поведения животных доминирующим стал эволюционный подход, который активно разрабатывался в российской науке К. Ф. Рулье, В. А. Вагнером, А. Н. Северцовым (Фабри, 1993; Филиппова, 2012). В дальнейшем инстинктивное поведение изучалось в плане его структуры и физиологических механизмов в работах этологов и физиологов ХХ в.: У. Крэга, К. Лоренца, Н. Тинбергена, Л. А. Орбели, И. П. Павлова и др. (Филиппова, 2012). В современных исследованиях анализируется уже исключительно поведенческий компонент инстинкта и его физиологическое обеспечение, основное внимание уделяется структуре инстинктивных действий (паттерны поведения, фиксированные последовательности действий) и физиологическим механизмам реакции на стимулы, которые вызывают инстинктивные действия. Актуальной остается связь инстинктивного поведения с другими формами поведения (научением, интеллектуальными или рассудочными действиями) в организации взаимодействия животного со средой.
Судьба же чувственного компонента инстинкта с XIX в. связана с психологией человека. Происхождение этого термина от латинского instinctus – «побуждение, стимул» – послужило основанием для применения его в объяснении влечений человека, которые расценивались как врожденные, естественные, иногда родственные животным или являющиеся инфантильными проявлениями. З. Фрейд, говоря о влечении, разводил понятие инстинкт (Instinkt) и влечение (Trieb) (Фрейд, 1989). Под инстинктом он понимал биологически наследуемое животное поведение, под влечением – психическое представительство соматического источника раздражения. Влечение, по Фрейду, возникает в результате актуализации потребности и направлено к объекту (это и есть объект влечения), при взаимодействии с которым удовлетворяется потребность. У человека есть врожденные влечения, первичными из которых являются либидо и мортидо. В позднейших психоаналитических текстах различение инстинктов и влечений несколько стерлось, и теперь они используются практически как синонимы. В психиатрии инстинкты также рассматриваются как влечения и присущие им поведенческие реакции, выделяются расстройства влечений – как чувственно-мотивационного, так и поведенческого их компонентов.
Подводя итог краткому экскурсу в историю изучения и трактовки инстинкта, можно заключить, что изначальная двухкомпонентная структура инстинкта, предложенная Хризиппом, подверглась дифференциации (что закономерно в развитии любого процесса, познания в том числе), и эти компоненты разошлись в разные науки: чувственный компонент – в психологию человека, поведенческий – в биологию поведения животных (этология, физиология). Строго говоря, современная зоопсихология все больше тяготеет к «объективному» – а на самом деле к биологическому методу исследования, а субъективный мир животного (что и является предметом психологии, независимо от того, какого носителя психики она изучает) до сих пор остается малодоступным. Попытки изучения этого субъективного мира животных, конечно, предпринимались неоднократно, но в основном со стороны содержания психического отражения: внутренняя структура интеллектуального поведения изучались в работах В. Келлера, Н. Н. Ладыгиной-Котс, Г. Г. Филипповой (Филиппова, 2012), отражение себя животными также было предметом исследования в конце XIX – начале ХХ столетий (И. А. Хватов, 2010). Эмоциональный компонент интерпретируется в теории привязанности (Дж. Боулби, 2003), хотя самих исследований привязанности на животных явно недостаточно. При этом психический компонент инстинкта остается в современной науке практически белым пятном. А в результате непонятно, что происходит с врожденными влечениями и действиями в онтогенезе человека и в его взрослом поведении. Я предлагаю посмотреть на эту проблему со стороны структуры инстинкта.
На самом деле, в инстинкте можно выделить не два, а три компонента: побуждающий компонент (влечение, основанное на чувственном переживании и мотивирующее активность субъекта по отношению к объекту влечения), поведение (исполнительная часть – что делать с объектом влечения) и переживание удовлетворения потребности, ради которого собственно все и происходит. В этологии этот «конец истории» (достижение биологического результата) был обозначен У. Крэйгом как консуматорный акт, являющийся завершающим в структуре инстинктивного поведения (Фабри, 1993). Анализируется консуматорный акт исключительно с позиции реализующих его действий животного. Но с психологической точки зрения именно этот момент и является тем, ради чего все поведение совершается и для чего появляется начальный компонент – влечение. Первый и третий из обозначенных выше структурных компонентов и представляют собой две взаимосвязанных части психического компонента инстинкта. С позиции двух современных подходов к анализу поведения, которое регулируется психическим отражением (теория функциональных систем и теория антиципации), существует непосредственная связь между влечением и переживанием удовлетворения потребности, и обе эти части отнесены непосредственно к субъекту, так как связаны с его потребностями. А поведенческий компонент является исполнительным звеном и отнесен к объекту, с которым субъект взаимодействует. В теории функциональных систем П. К. Анохина конечный компонент – это акцептор результата действия, который представлен как чувственное переживание, и его достижение регулирует работу функциональной системы по принципу обратной связи (Анохин, 1980). Обратная связь основана на предвосхищении (антиципации) результата. А это уже чисто психическое образование.
Теперь следует задать вопрос: что из этих компонентов является врожденным и как эта трехчленная структура преобразуется в фило- и онтогенезе. Для этого представим инстинкт как функциональную систему: есть пусковой стимул, который попадает на систему опознания (в данном случае это врожденная нервная модель стимула) – в результате действия стимула возникает влечение, существует программа поведения (при инстинктивном поведении – врожденный фиксированный паттерн действия) и акцептор результата действия (образ цели, в данном случае – врожденная модель переживания удовлетворения потребности). Акцептор результата действия связан с программой поведения системой обратной афферентации. При достижении результата происходит, по Н. Тинбергену, переключение на следующую в цепочке инстинктивных актов нервную модель стимула (Тинберген, 1985). И пока актуальная функциональная система не завершится достижением результата, переключение невозможно. Такая схема позволяет понять, в каком месте заложена пластичность инстинктивного акта поведения и что происходит при трансформации инстинкта в фило- и онтогенезе.
При полностью инстинктивной регуляции деятельности все три компонента являются врожденными и жестко связанными в единую последовательность. В филогенезе с усложнением связей организма со средой происходит увеличение количества потребностей (во взаимосвязи начального и конечного компонентов и соотвествующих начальному компоненту нервных моделей стимулов) и усложнение поведенческого компонента. Пластичность инстинкта в первую очередь связана с изменением среднего компонента, что происходит за счет включения научения и впоследствии интеллекта, а у человека – сознания. Соответственно, усложняется и весь механизм регуляции исполнительного звена. В принципе, до определенных стадий филогенеза этим все и исчерпывается. Набор потребностей достаточно неизменен и, можно сказать, инстинктивен. Хотя, конечно, изменяется и варьируется опознание стимула. Тут большое поле для включения научения и более сложных форм регуляции деятельности. Объекты влечения могут осваиваться прижизненно, и нервные модели стимулов тоже строятся прижизненно. Появляется онтогенез – как прижизненное соединение инстинктивных и приобретенных компонентов взаимодействия субъекта со средой. На основе импринтинга и облигатного научения появляется образ объекта, в исследовательской деятельности и позднее в игре происходит «переподключение» исполнительного звена к разным парам «влечение – удовлетворение потребности».
В дальнейшем, на высших стадиях филогенеза (на стадии интеллекта и сознания) появляется прижизненное формирование новых пар «влечение – удовлетворение потребности». Это может происходить с конца функциональной системы – случайное появление новых переживаний удовлетворения становится желаемым, это переходит во влечение получать эти переживания с помощью определенного поведения. А возможно узнавание о потребностях от других особей (что характерно для человека) и организация всей системы целенаправленно, до выстраивания новой полноценной функциональной системы. Это уже практически совсем выходит за рамки инстинкта, остается только внутренний нейрогуморальный механизм, обеспечивающий подкрепление в форме удовольствия (Филиппова, 1998, 2012).
Таким образом, в филогенезе и онтогенезе высших животных и человека «раздвигаются рамки» инстинкта, и его структура наполняется и изменяется за счет других форм регуляции деятельности, т. е. изменяется не инстинкт, а взаимодействие субъекта со средой, в которое включаются другие регуляторные механизмы. В этом случае просто невозможно говорить об инстинктах у человека – у него их просто нет, а есть только отдельные врожденные компоненты, объединяющиеся в адаптивное поведение прижизненно с помощью импринтингов, облигатного и факультативного научения, игры и т. д. Это могут быть врожденные – инстинктивные – влечения, отдельные нервные модели стимулов (в основном для безусловных рефлексов или формируемые пренатально), модели переживания удовлетворения потребностей (акцепторы результата действия) и сложный онтогенетический механизм формирования всех связей с миром. Для примера можно описать формирование материнства в онтогенезе (Филиппова, 1998).
Пресловутый «материнский инстинкт» трактуется как чувство матери к ребенку (положительно-эмоциональное отношение, умиление, тревога при его дискомфорте), способность понимать состояние ребенка и стремление заботиться о нем. Из всего этого к инстинктивным компонентам можно отнести реакцию на ключевые стимулы от младенца (плач, внешний вид, запах), которая, как показывает практика и исследования, изначально даже не определена эмоционально. В ситуации стресса, неготовности к материнству, при отсутствии или искажении индивидуального опыта чувство к ребенку может быть отрицательным: агрессия, раздражение, страх, тревога, отвращение. Положительно-эмоциональное отношение и стремление к контакту с ребенком возникает у женщины в процессе общения с матерью и другими близкими взрослыми в раннем возрасте, в игровой деятельности и опыте общения с младенцами в детстве и отрочестве. Запуску материнских чувств способствует процесс родов и встреча с новорожденным, когда создаются эндогенные условия для запечатления и проявления влечения к ребенку. Однако даже при всех прочих положительных событиях (планируемая и хорошо протекающая беременность и физиологические роды с рождением здорового ребенка) вместо ожидаемых чувств может возникнуть растерянность, тревога и даже депрессия, которые могут быть связаны с отсутствием опыта заботы о ребенке и внешней поддержки и помощи. В данном случае на основе весьма фрагментарных врожденных компонентов прижизненно формируются все части функциональной системы: влечение, удовлетворение от деятельности, исполнительные звенья (Филиппова, 2015).