



Павел Симонов
Эмоциональный мозг
Заметим, что стадия прагматической неопределенности имеет место не только при формировании сложной системы условных реакций – динамического стереотипа, но и при формировании одиночного условного рефлекса, который также представляет систему, состоящую из двух или нескольких безусловных [Асратян, 1953, с. 208]. Еще в 1924 г. советский психиатр В. П. Осипов проницательно назвал первую стадию образования любого условного рефлекса – стадию генерализации – «эмоциональной» в отличие от более поздней «интеллектуальной, познавательной» стадии хорошо упроченного рефлекса [Осипов, 1924]. Эти две стадии (динамику ослабления, а затем и постепенного исчезновения эмоционального напряжения) легко продемонстрировать на примере выработки оборонительного условного рефлекса у человека. Испытуемых просили нажимать на ключ через 20 с после короткого звукового сигнала (рис. 1). Если субъект нажимал на ключ раньше 19 с после подачи сигнала или позднее 21 с, ему наносили на кожу предплечья болевое раздражение электрическим током порядка 60–90 В. После каждой пробы субъекта информировали о времени его реакции. Степень эмоционального напряжения измеряли по изменению частоты сердцебиений. Одновременно регистрировали кожногальванический рефлекс. Нарастание частоты сердцебиений оценивали по суммарной продолжительности первых трех ударов сердца после звукового сигнала и последних трех ударов перед двигательной реакцией (скорость движения бумаги – 1,5 см/с). На протяжении первых 10 предъявлений условного сигнала субъект знал, что раздражения током не будет. Затем несколько раз применяли изолированное электрическое раздражение для того, чтобы определить интенсивность тока, не менее чем в три раза превышающую болевой порог. Эта интенсивность сохранялась постоянной на протяжении всего эксперимента.
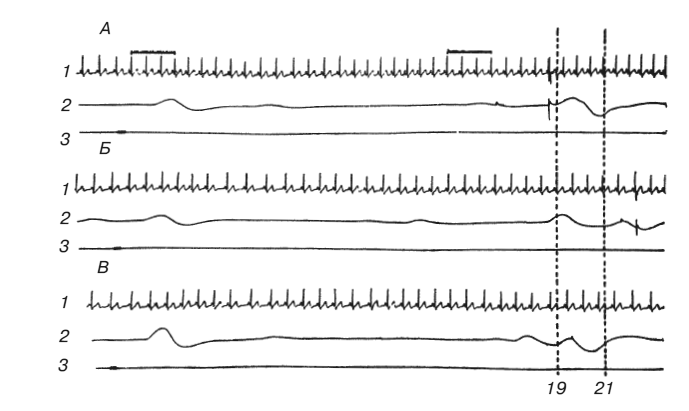
Рис. 1. Условный оборонительный рефлекс у человека
А, Б – ошибочные реакции, сопровождавшиеся болевым раздражением током; В – правильная реакция; 1 – ЭКГ; 2 – КГР, отметка реакции и тока; 3 – звуковой сигнал. Прерывистыми линиями отмечена «зона безопасности»
На рис. 2 сплошная кривая показывает время двигательных реакций в последовательных пробах. Раздражения током наносились, когда ошибки субъекта превышали допустимое отклонение (обозначено двумя горизонтальными линиями). Прерывистая линия показывает изменения продолжительности шести сердечных сокращений в пересчете на число ударов в минуту. Верхняя горизонтальная линия обозначает среднюю частоту пульса в фоне до начала выработки условного рефлекса.
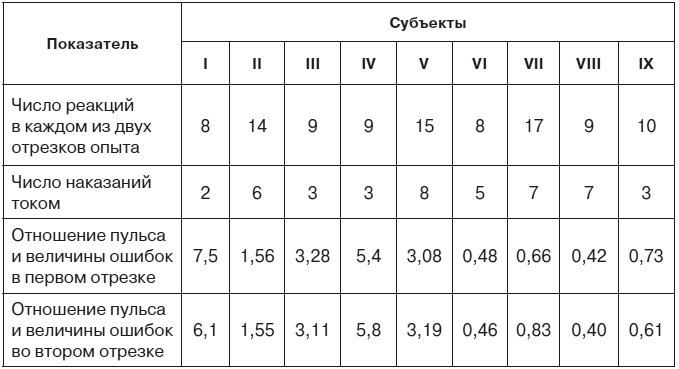
Если сравнить две стадии эксперимента, содержащие одинаковое число проб и одинаковое количество болевых раздражений током, можно убедиться, что суммарное отклонение частоты сердцебиений от исходного фона в этих двух частях эксперимента различно. Следовательно, оно зависит не только от количества наказаний. Опыты на 9 испытуемых показали, что суммарное изменение частоты сердцебиений пропорционально суммарному отклонению времени двигательных реакций от заданной величины, то есть пропорционально степени совершенства, точности и надежности условного оборонительного рефлекса (табл. 1). Это правило справедливо и для тех случаев, где величина ошибок (но не их количество, не количество болевых раздражений) нарастала вместе с нарастанием частоты сердцебиений и где, следовательно, динамику вегетативных сдвигов нельзя было объяснить привыканием к болевым стимулам по мере их повторения.
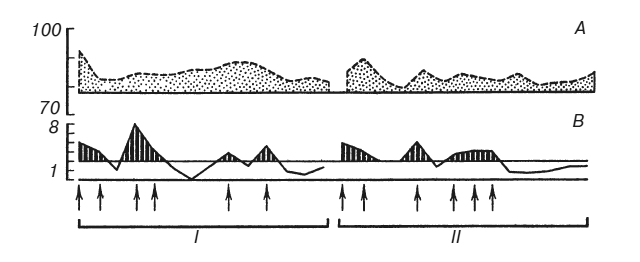
Рис. 2. Выработка условного оборонительного рефлекса
А — нарастание частоты сердцебиений по сравнению с фоном (уд/мин); В – отклонение времени двигательных реакций от заданной величины (с). Абсцисса – последовательные пробы на протяжении двух стадий эксперимента I и II. Стрелки – раздражения током
Таблица 1. Отношение суммарного изменения частоты сердцебиений к суммарной величине ошибок в опытах с девятью исследуемыми лицами
Эксперименты на животных также показывают, что мозг прогнозирует вероятность наказания в зависимости от степени совершенства инструментального двигательного рефлекса. Л. А. Преображенская вырабатывала условный оборонительный рефлекс у собак в ситуации, где подъем передней лапы до определенного уровня и удержание ее на этом уровне в течение 10 с предотвращали болевое раздражение током противоположной задней лапы (методика Г. П. Зеленого, Г. В. Скипина, Р. Л. Винник и др.). Условный звуковой сигнал подавали за 10 с до болевого раздражения. Сочетание звука с болевым раздражением до выработки условного двигательного рефлекса вело к нарастанию амплитуды и процентного содержания тета-ритма в частотном спектре электрической активности дорзального гиппокампа (рис. 3). Количественный анализ обнаружил позитивную корреляцию изменений суммарного напряжения гиппокампального тета-ритма (измеренного по показаниям интегратора) с частотой сердцебиений. Оба симптома заметно ослабевали по мере стабилизации двигательного навыка, надежно избавляющего животное от боли. Любые затруднения в осуществлении движений вели к повторному нарастанию тета-ритма. Таким образом, опыты Л. А. Преображенской [1969; см. также: Konorski, Santibanez, Beck, 1968] показали, что интенсивность гиппокампального тета-ритма зависит не от двигательной активности самой по себе, но от эффективности двигательных актов, от их влияния на вероятность предотвращения болевых раздражений. Вместе с тем на степень эмоционального напряжения оказывает свое влияние и потребностно-мотивационный фактор: в опытах с оборонительными условными рефлексами суммарное напряжение тета-ритма и учащение сердцебиений были выражены значительно сильнее, чем в опытах с пищевыми.
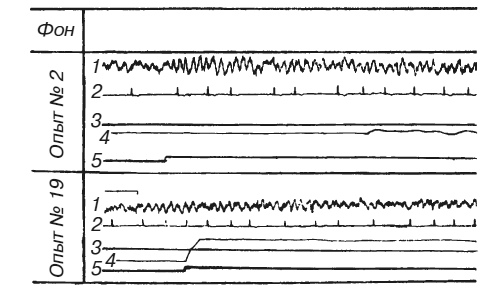
Рис. 3. Изменения гиппокампального тета-ритма и частоты сердцебиений во время выработки инструментального условного оборонительного рефлекса у собаки
1 – ЭГ дорзального гиппокампа; 2 – ЭКГ; 3 – уровень выключения тока; 4 – подъемы передней лапы, выключавшей ток; 5 – условный звуковой сигнал (по Л. А. Преображенской)
Заканчивая обзор данных, свидетельствующих об исчезновении эмоционального напряжения по мере выработки адекватной условной реакции, П. Фресс пишет: «Прежде всего следует подчеркнуть, что не существует эмоциогенной ситуации как таковой. Она зависит от отношения между мотивацией и возможностями субъекта» [Фресс, 1975, с. 133]. Предположение о том, что даже хорошо упроченный инструментальный оборонительный условный рефлекс продолжает мотивироваться страхом, как бы замещающим потребность избегания боли, подвергнуто аргументированной критике Ж. Кюттеном: «…потребность избегания болезненного стимула продолжает действовать (до тех пор, пока животное не усвоит, что сигнал не предвещает опасности), но поскольку животное не испытывает ни боли от стимула, ни угрозы опасной ситуации, оно не проявляет более эмоциональной реакции страха» [Кюттен, 1975, с. 70].
Литература переполнена экспериментальными данными, свидетельствующими о зависимости эмоционального напряжения от величины потребности (мотивации) и прогнозирования вероятности ее удовлетворения. Например, было установлено, что частота пульса у банковских служащих зависит от степени их ответственности (счет банкнотов различного достоинства) и количества информации, содержащейся в одной операции [Gantchev, Danev, Kitcheva, 1967]. Эмоциональные реакции обезьян закономерно зависят от изменения вероятности пищевого подкрепления [Meiges, Popper, 1976]. По данным Д. Н. Меницкого и М. М. Хананашвили [1969], наибольшее эмоциональное напряжение у собак (визг, лай, чесание, царапанье кормушки) наблюдалось при вероятности подкрепления 1:4, а по мере продолжения опыта – при 1:2. Значение информационного фактора выступает особенно отчетливо в опытах со спаренными животными, когда оба партнера получают равное количество ударов током, но только один из них может предотвратить наказание соответствующей инструментальной реакцией. Показано, что именно у этого животного постепенно исчезают признаки страха, предотвращается изъявление слизистой желудка и кишечника [Desiderato, Newman, 1971; Jonas, Jonas, 1975; Starr, Mineka, 1977]. Мотивационный и информационный факторы генеза эмоционального напряжения имеют различный вес у разных животных одного и того же вида. По данным Л. А. Преображенской [1974], максимальное учащение сердечного ритма наблюдалось у одних собак при 30 %-ном пищевом подкреплении, а у других – при 5 %-ном. Следовательно, для собак второй группы суммарный «проигрыш» в удовлетворении пищевой потребности имел большее значение, чем неопределенность экспериментальной ситуации.
Имеются сведения и о том, что две составляющие эмоционального напряжения по-разному сказываются на величине различных вегетативных сдвигов. В опытах с участием человека величина побуждения (размер платы за правильное решение) преимущественно влияла на частоту пульса, дыхания и уровень электрического сопротивления кожи, а трудность задачи (количество выборов) – на объемный пульс и кожногальванический рефлекс [Wilkinson, El-Beheri, Gieseking, 1972]. Преимущественная связь кожногальванического рефлекса с информационным фактором обнаружена и в опытах, где кожногальванический рефлекс был слабее при ожидании болевого раздражения током с высокой вероятностью, чем при более редких, но трудно прогнозируемых ударах [Epstein, Bahm, 1971].
Отражательно-оценочная функция эмоций
«Первые понятия, с которых начинается какая-нибудь наука, – писал Н. И. Лобачевский, – должны быть ясны и приведены к самому меньшему числу Тогда только они могут служить прочным и достаточным основанием учения» [Лобачевский, 1976, с. 39]. Суммируя результаты собственных опытов и данные литературы, мы пришли в 1964 г. к выводу о том, что эмоция есть отражение мозгом человека и животных какой-либо актуальной потребности (ее качества и величины) и вероятности (возможности) ее удовлетворения, которую мозг оценивает на основе генетического и ранее приобретенного индивидуального опыта.
В самом общем виде правило возникновения эмоций можно представить в виде структурной формулы:
Э = f [П, (Ин – Ис), …],
где Э — эмоция, ее степень, качество и знак; П — сила и качество актуальной потребности; (Ин – Иc) – оценка вероятности (возможности) удовлетворения потребности на основе врожденного и онтогенетического опыта; Ин — информация о средствах, прогностически необходимых для удовлетворения потребности; Ис — информация о средствах, которыми располагает субъект в данный момент.
Разумеется, эмоция зависит и от ряда других факторов, одни из которых нам хорошо известны, а о существовании других мы, возможно, еще и не подозреваем. К числу известных относятся:
• индивидуальные (типологические) особенности субъекта, прежде всего индивидуальные особенности его эмоциональности, мотивационной сферы, волевых качеств и т. п.;
• фактор времени, в зависимости от которого эмоциональная реакция приобретает характер стремительно развивающегося аффекта или настроения, сохраняющегося часами, днями и неделями;
• качественные особенности потребности. Так, эмоции, возникающие на базе социальных и духовных потребностей, принято именовать чувствами. Низкая вероятность избегания нежелательного воздействия породит у субъекта тревогу, а низкая вероятность достижения желаемой цели – фрустрацию и т. д. и т. п.
Но все перечисленные и подобные им факторы обусловливают лишь вариации бесконечного многообразия эмоций, в то время как необходимыми и достаточными являются два, только два, всегда и только два фактора: потребность и вероятность (возможность) ее удовлетворения.
Во избежание недоразумений (о них мы специально будем говорить в следующей главе) остановимся на уточнении употребляемых нами понятий. Термин «информация» мы используем, имея в виду ее прагматическое значение, то есть изменение вероятности достижения цели (удовлетворения потребности) благодаря получению данного сообщения [Харкевич, 1960]. Таким образом, речь идет не об информации, актуализирующей потребность (например, о возникшей опасности), но об информации, необходимой для удовлетворения потребности (например, о том, как эту опасность избежать). Под информацией мы понимаем отражение всей совокупности средств достижения цели: знания, которыми располагает субъект, совершенство его навыков, энергетические ресурсы организма, время, достаточное или недостаточное для организации соответствующих действий, и т. п. Спрашивается, стоит ли в таком случае пользоваться термином «информация»? Мы полагаем, что стоит, и вот почему. Во-первых, мозг, генерирующий эмоции, имеет дело не с самими навыками (куда входит и тренировка периферического исполнительного аппарата), не с самими энергетическими ресурсами организма и т. д., а с афферентацией из внешней и внутренней среды организма, то есть с информацией об имеющихся средствах. Во-вторых, все многообразие сведений о необходимом для удовлетворения возникшей потребности и реально имеющемся в данный момент у субъекта трансформируется мозгом в единый интегральный показатель – в оценку вероятности достижения цели (удовлетворения потребности). Оценка же вероятности по самой природе своей есть категория информационная.
Термин «потребность» мы употребляем в его широком Марксовом понимании, отнюдь не сводимом к одному лишь сохранению (выживанию) особи и вида. «Дай человеку то лишь, без чего не может жить он, – ты его сравняешь с животным», – писал Шекспир в «Короле Лире», но и потребности животных не ограничиваются самосохранением. Подробнее проблема потребностей будет рассмотрена нами ниже, а сейчас мы ограничимся лишь возможно более точным определением термина. Нередко потребность квалифицируют как нужду в чем-либо, но подобное определение есть не более чем игра в синонимы. По нашему мнению, потребность есть избирательная зависимость живых организмов от факторов внешней среды, существенных для самосохранения и саморазвития, источник активности живых систем, побуждение и цель их поведения в окружающем мире. Соответственно поведение мы определим как такую форму жизнедеятельности, которая может изменить вероятность и продолжительность контакта с внешним объектом, способным удовлетворить имеющуюся у организма потребность.
По справедливому замечанию В. Ф. Сержантова и Т. И. Сержантовой [1976, с. 14], именно категория потребности способна сыграть роль «концептуального моста», соединяющего естественнонаучное знание, резюмированное в понятии «организм», и психологическое и социальное знание о природе человека, обобщенное в понятии «личность». Согласны мы и с Г. Г. Дилигенским в том, что «функцию фиксации определенной направленности различных потребностей, способов действия по их удовлетворению выполняют установки, ценности, интересы, цели субъекта. В марксистской психологической литературе все эти уровни мотивации обычно характеризуются как производные от потребностей» [Дилигенский, 1977, с. 115].
Выявление потребности, инициировавшей тот или иной поступок, представляет наиболее трудную задачу и для теоретического анализа, и для практики воспитания. «За тридцать лет работы в школе, – писал В. А. Сухомлинский, – мне довелось разбирать сто как будто бы совершенно одинаковых поступков: подросток скрыл от родителей неудовлетворительные оценки, поставленные учителем. Но в каждом случае есть свои причины, свои нравственные и эмоциональные мотивы…» [Сухомлинский, 1979, с. 49]. Эвристическая ценность эмоций как раз и состоит в том, что в отличие от поступка они могут служить одним из наиболее тонких и объективных индикаторов интересующей нас потребности.
К понятию «потребность» наиболее тесно примыкает феномен мотивации. Хорошее представление об истории изучения мотивации дает коллекция статей, собранная В. А. Расселлом [Russell, 1970]. Мотивация представляет второй этап организации целенаправленного поведения по сравнению с актуализацией потребности (рис. 4), ее можно рассматривать как «опредмеченную потребность». Не существует мотивации без потребностей, но вполне возможно встретить потребность, не ставшую мотивацией. Так, человек может испытывать острейшую потребность в витаминах и не быть мотивированным, поскольку он не знает о причине своего состояния. Собака, лишенная коры больших полушарий головного мозга, под влиянием голода (потребности в пище) приходит в состояние сильнейшего двигательного возбуждения. Тем не менее говорить о пищевой мотивации здесь нельзя, поскольку собака не прикасается к пище, лежащей у нее под ногами. Итак, мотивация есть физиологический механизм активирования хранящихся в памяти следов (энграмм) тех внешних объектов, которые способны удовлетворить имеющуюся у организма потребность, и тех действий, которые способны привести к ее удовлетворению. При таком понимании нейрофизиологии мотивации нам, естественно, трудно согласиться с представлением об исходном мотивационном возбуждении структур гипоталамуса. Только вовлечение гипоталамуса в условнорефлекторную деятельность (о чем речь пойдет ниже) придает активности гипоталамических структур мотивационный характер. В последние годы Э. А. Асратян представил стройную систему теоретических и экспериментальных аргументов в пользу концепции, рассматривающей физиологические механизмы мотивации как результат взаимодействия сложнейших жизненно важных безусловных рефлексов с прямыми и обратными условными связями. Особое значение для понимания механизмов мотивации имеют обратные условные связи, в том числе их тонический вариант, благодаря которому происходит избирательное повышение возбудимости структур, воспринимающих условный сигнал возможного подкрепления. Рассматривая проблему мотивации поведения в свете рефлекторной теории, Э. А. Асратян подчеркнул: «…активация обратных условных связей может иметь своим следствием не только условнорефлекторное воспроизведение рефлекса первого в сочетанной паре раздражителя, но и может ограничиться одним лишь повышением возбудимости центральных структур последнего» [Асратян, 1974, с. 14].
Вернемся к анализу следствий, вытекающих из «формулы эмоций». Низкая вероятность удовлетворения потребности (Ин больше, чем Ис) ведет к возникновению отрицательных эмоций. Возрастание вероятности удовлетворения по сравнению с ранее имевшимся прогнозом (Ис больше, чем Ин) порождает положительные эмоции. Примером последнего случая могут служить эмоциональные реакции, возникающие у человека в процессе решения задачи по дифференцированию зрительных сигналов.
В наших опытах на экране, установленном перед испытуемым, проецировались наборы из пяти цифр – единиц и нулей. Испытуемого предупреждали, что некоторые из кадров, содержащие общий признак (например, два нуля подряд), будут сопровождаться гудком. Задача испытуемого состояла в обнаружении этого общего признака. В ходе опыта испытуемый должен был через микрофон сообщать экспериментатору о возникающих у него предположениях. Время экспозиции каждого кадра составляло 10 с. При этом регистрировали два отведения электроэнцефалограммы, электрокардиограмму и кожногальванический рефлекс (КГР). Прежде чем испытуемый получал инструкцию, производили угашение КГР на смену кадров и гудок.
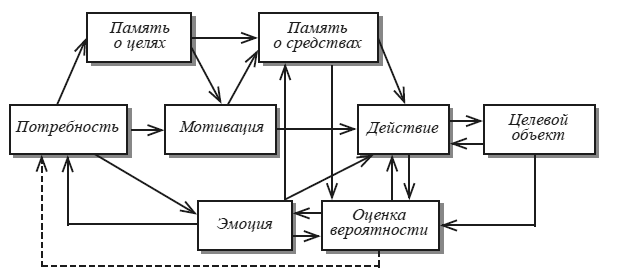
Рис. 4. Потребность, мотивация и эмоция в структуре целостного поведенческого акта Пунктир – тормозящее влияние.
Подобная задача является типичным примером творческой интеллектуальной деятельности, где сформулированы «требования задачи», но отсутствует «искомое», которое только еще предстоит найти [Брушлинский, 1966]. До возникновения первой (как правило, ошибочной) гипотезы относительно подкрепляемого признака ни новые кадры, ни гудок не вызывали КГР (рис. 5, Д). Этот факт лишний раз свидетельствует о том, что ориентировочной реакции на «новый раздражитель» не существует в природе. Ориентировочная реакция есть реакция на результат сравнения внешнего стимула с «нервной моделью» [Соколов, 1960] в самом широком ее понимании от энграммы конкретного стимула до обобщенной модели всей ситуации, включая ее возможные изменения, прогнозируемые субъектом. Возникновение гипотезы сопровождается КГР (рис. 5, Б). Поскольку КГР имеет сравнительно большой латентный период, рисунок ясно показывает, что колебание кожных потенциалов связано с моментом осознания гипотезы, а не с речевым сообщением испытуемого. После формирования гипотезы возможны две ситуации, которые мы рассматриваем в качестве экспериментальных моделей отрицательной и положительной эмоциональных реакций.
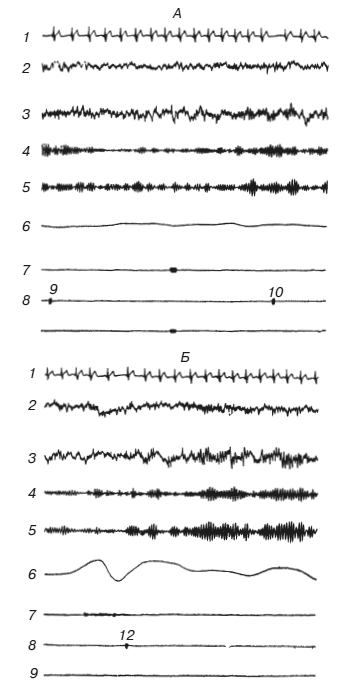
Рис. 5 (А, Б). КГР до возникновения гипотезы о подкрепляемом признаке (А), в момент возникновения гипотезы (Б), при рассогласовании (В) и в случае подтверждения гипотезы (Г)
1 — ЭКГ; 2, 3 — ЭЭГ; 4, 5 — анализ альфа-ритма ЭЭГ; 6 – КГР; 7 – отметка речевого ответа испытуемого; 8 – отметка смены кадров; 9 – отметка гудка. Цифры на 8 – номера кадров.

Рис. 5 (В, Г)
1 – ЭКГ; 2, 3 — ЭЭГ; 4, 5 — анализ альфа-ритма ЭЭГ; 6 – КГР; 7 – отметка речевого ответа испытуемого; 8 – отметка смены кадров; 9 – отметка гудка. Цифры на 8 – номера кадров.
Первый случай представлен на рис. 5, В. Гипотеза неверна, и кадр 25, содержащий подкрепляемый признак, не вызывает КГР. Когда же гудок показывает испытуемому, что он ошибся, регистрируется КГР как результат рассогласования гипотезы с наличным раздражителем – случай, предусмотренный концепциями «акцептора результата действия» П. К. Анохина, «нервной модели стимула» Е. Н. Соколова и им подобными. Испытуемый несколько раз меняет гипотезу, и в какой-то момент она начинает соответствовать действительности (рис. 5, Г). Теперь уже само появление подкрепляемого кадра 30 вызывает КГР, а его подкрепление гудком приводит к еще более сильным кожногальваническим сдвигам.
Как понять подобный эффект? Ведь в данном случае произошло полное совпадение гипотезы («акцептора результата действия», «нервной модели» и т. д.) с наличным стимулом. Отсутствие рассогласования должно было бы повлечь за собой отсутствие КГР и других вегетативных сдвигов. На самом деле в последнем случае мы также встречаемся с рассогласованием, но рассогласованием иного рода, чем при проверке ложной гипотезы. Формирующийся в процессе повторных сочетаний прогноз содержит не только афферентную модель цели, не только ее семантику, но и вероятность достижения этой цели. В момент подкрепления кадра 30 гудком прогнозируемая вероятность решения задачи (правильности гипотезы) резко возросла, и это рассогласование прогноза с поступившей информацией привело к сильной КГР как вегетативному компоненту положительной эмоциональной реакции. Значение вероятностного прогнозирования для генеза положительных эмоций описано И. М. Фейгенбергом [1965] при исследовании нарушений аффективной сферы у больных шизофренией.
Фактор позитивного рассогласования обнаруживается и в сфере эстетики. В. М. Волькенштейн [1931] определил прекрасное как «целесообразное и сложное (трудное) преодоление». «Шахматная красота возникает там, где есть целесообразность ходов, связанная с перипетией, то есть неожиданным парадоксальным (на первый взгляд) преодолением в затруднительном положении. Таковы моменты пожертвования. Если же победа дается постепенным накоплением мелких преимуществ, без увлекательной комбинации, ведущих через выигрыш пешки или качества к верному выигрышу, такая игра может быть названа солидной, выдержанной, даже поучительной – даже тонкой; но красивой ее назвать нельзя» [Волькенштейн, 1931, с. 45–46]. «Хорошие дни выпадают на долю разумных людей, – пишет Сигрид Унсет, – но лучшие дни достаются тому, кто посмеет быть безумным».
Информационная теория эмоций справедлива не только для сравнительно сложных поведенческих и психических актов, но для генеза любого эмоционального состояния. Например, положительная эмоция при еде возникает за счет интеграции голодового возбуждения (потребность) с афферентацией из полости рта, свидетельствующей о растущей вероятности удовлетворения данной потребности. При ином состоянии потребности та же афферентация окажется эмоционально безразличной или генерирует чувство отвращения.
До сих пор мы говорили об отражательной функции эмоций, которая совпадает с их оценочной функцией. Обратите внимание, что цена в самом общем смысле этого понятия всегда есть функция двух факторов: спроса (потребности) и предложения (возможности эту потребность удовлетворить). Но категория ценности и функция оценивания становятся ненужными, если отсутствует необходимость сравнения, обмена, то есть необходимость сопоставления ценностей. Вот почему функция эмоций не сводится к простому сигнализированию воздействий полезных или вредных для организма, как полагают сторонники «биологической теории эмоций». Воспользуемся примером, который приводит П. К. Анохин [1964, с. 342]. При повреждении сустава чувство боли ограничивает двигательную активность конечности, способствуя репаративным процессам. В этом интегральном сигнализировании «вредности» П. К. Анохин видел приспособительное значение боли. Однако аналогичную роль мог бы играть механизм, автоматически, без участия эмоций тормозящий движения, вредные для поврежденного органа. Чувство боли оказывается более пластичным механизмом: когда потребность в движении становится очень велика (например, при угрозе самому существованию субъекта), движение осуществляется, невзирая на боль. Иными словами, эмоции выступают в роли своеобразной «валюты мозга» – универсальной меры ценностей, а не простого эквивалента, функционирующего по принципу: вредно – неприятно, полезно – приятно, как об этом принято писать в десятках дублирующих друг друга статей.
Из существа отражательно-оценочной функции эмоций вытекают их регуляторные функции. Мы начнем их анализ с функций, регулирующих индивидуальное поведение.
