



Роб Десалл
Чувства: Нейробиология сенсорного восприятия
1.3 Бактериальное чувство кворума
Микробам часто необходимо чувствовать плотность популяции, чтобы реагировать на изменения окружающей среды. Классический пример этой способности – чувство кворума, присущее биолюминесцентным бактериям Aliivibrio fischeri, населяющим световой орган гавайского короткохвостого кальмара (Euprymna scolopes). Между кальмаром и бактериями установились мутуалистические (взаимовыгодные) отношения: кальмар взращивает в себе A. fischeri, а бактерии, в свою очередь, освещают фотогенный орган кальмара, используемый им для маскировки от хищников. Однако бактериям важно понимать, когда именно надо «зажечь свет»: «светить всегда, светить везде» – ужасное расточительство энергии. Поэтому в световом органе кальмара развился механизм регуляции экспрессии белков, продуцирующих биолюминесценцию, который использует хитрую способность бактерий ощущать размер своей популяции. Бактерии производят белок, называемый индуктором, который они идентифицируют благодаря наличию у них другого белка – рецептора. Когда рецептор и индуктор связываются друг с другом при помощи механизма «замок – ключ», происходит биолюминесцентная реакция, поскольку в бактериальном геноме постепенно «включается» цепочка генов. Когда в световом органе находится недостаточное количество A. fischeri, концентрация индуктора настолько низка, что тот не может эффективно связаться с рецептором и свет не генерируется. Такой вид восприятия происходит полностью на молекулярном уровне.
Во-первых, плотная кластеризация генов, участвующих в формировании магнитосом и содержащих их органелл, указывает на единый механизм этого феномена (по крайней мере у типов, где обнаружены магнитотактические бактерии). Во-вторых, эти гены расположены на так называемом колонизационном участке ДНК, который может передвигаться по горизонтали к другим видам. Этот горизонтальный перенос способен осуществить и ускорить эволюцию магнитотактического признака у других бактерий.
Если никакое другое магнитное поле не мешает, бактерии выравниваются по магнитному полю Земли. Почему микробы делают это? Для выживания им необходимы питательные вещества, а зная направление магнитного поля, легче отыскать подходящую питательную среду. Например, микробы, относящиеся к типам магнитотактических бактерий, жаждут оказаться в тепленьком местечке – в благотворной для них среде: бескислородной зоне или зоне с низким содержанием кислорода. Они эволюционировали в подобной среде и теперь при помощи сильных флагелл (жгутиков) перемещаются в ее поисках. Оказывается, из-за искривления Земли ее магнитное поле направлено не только с севера на юг, но и под углом к ее поверхности. И знание этого угла позволяет бактериям ориентироваться над и под землей. Способность чувствовать магнитное поле помогает магнитотактическим бактериям добраться к бескислородным зонам, находящимся глубоко в недрах, наиболее эффективным путем.
Что же происходит, когда микробы ошибочно реагируют не на магнитное поле Земли, а на какое-то другое? Исследователи Корейского института науки и техники создали крошечный аппарат в виде чашки Петри на платформе, под которой образуется магнитное поле. Магнитное устройство контролируемо вращается таким образом, что преодолевает магнитное поле Земли и управляет поведением бактерий. Магнитотактические бактерии помещаются в чашку Петри, джойстик заводит магнитное поле – и под ритм «Хлопкоглазого Джо»[3] бактерии пускаются в пляс: в лаборатории начинается линейный танец[4]. Смотреть, как танцуют одноклеточные организмы – а они довольно неплохо это делают и демонстрируют достойную версию движений танца в линию, вращаясь в унисон слева направо и двигаясь вперед-назад, – весьма унизительно для такого плохого танцора, как я. Танец – это только метафора, конечно, и очень важно понять, что же именно стоит за ней.
Отдельные клетки должны знать, где они находятся в пространстве, с чем они вступают в контакт и когда именно. Поскольку миллиарды лет назад солнечный свет был преобладающим фактором воздействия на живые организмы, некоторым бактериям необходимо было знать, где тот находится, чтобы использовать его как средство выживания. Поэтому отдельные клетки научились обнаруживать внешние факторы, такие как гравитация, свет и содержащиеся в окружающей среде химические вещества, довольно сложными и эффективными способами. Андрий Анишкин и его коллеги предположили, что тактильное восприятие в оригинальном смысле (как они его называют) служит хорошим аргументом в пользу того, что это первое и, возможно, самое важное чувство, которым наделена клетка. Но все, что касается порядка, в котором клетки и организмы развивали другие чувства, может быть только предположением. Хотя мы и можем придумать довольно логичные объяснения, почему и как клетки формировали тот или иной способ восприятия.
Для некоторых бактерий, как и для растений, свет – это «еда». Цианобактерии, или синезеленые водоросли, способны к фотосинтезу. Молекулы взаимодействуют со светом, поглощая фотоны, и именно за счет этого бактерии и выживают. Механизмы использования света в качестве пищи у растений и бактерий практически одинаковые. На первый взгляд это выглядит полной чушью, ведь бактерии и растения тесно не связаны, у них нет явных общих предков или потомков, что объяснило бы возможное наличие у растений характеристик, присутствующих у бактерий. Но все это лишь до тех пор, пока мы не рассмотрим происхождение хлоропласта – органеллы, превращающей свет в питательное вещество для клеток растений. По сути хлоропласт – это наследие древних цианобактерий, поглощенных клетками предшественников современных растений. Симбиоз, вызванный поглощенными цианобактериями, настолько хорошо сказался на жизни древней растительной клетки, что она застряла в эволюционном контексте и теперь представляет собой основу растительной жизни на планете. История того, как древние эукариотические клетки поглощали различные виды бактерий, сложна и иногда запутанна. Некоторые растительные клетки многократно поглощали другие клетки, и даже многократно поглощенные клетки поглощались снова и снова.
Другой способ использования света бактериями – это изменение молекулярных свойств класса молекул, называемых опсинами. Эти молекулы встроены в клеточные мембраны, где фотоны могут натолкнуться на них. Во внутренней структуре опсинов заперты еще более мелкие молекулы – хромофоры. Хромофор, цепляющийся за внутренности опсина, приводит бо́льшую молекулу, встроенную в клеточную мембрану, в специфическое неактивное состояние. Когда свет определенной длины волны попадает в клетку, он также натыкается на хромофор и вызывает его смещение, а структура самого опсина изменяется, вызывая другие реакции в клетке.
У некоторых одноклеточных бактерий в клеточную мембрану встроена молекула родопсина, которая реагирует со светом. Но в отличие от более сложных организмов родопсин у бактерий действует как насос, который доставляет высокие концентрации ионов хлора или перемещает протоны в клетку, что, в свою очередь, изменяет способ дальнейшей жизнедеятельности клетки. У одноклеточных эукариот тоже есть родопсины, которые реагируют на попадание света. Родопсин бактерий довольно сильно отличается от опсинов высших эукариотов, поэтому неизвестно, являются ли опсины позвоночных продуктом его эволюции. Дело в том, что механизмы обнаружения света опсинами и родопсинами схожи и дают представление о том, как высшие животные воспринимают свет. Еще одна особенность заключается в том, что у одноклеточных организмов эти механизмы реализуются с помощью белков без централизованной обработки информации в мозге. «Решения», которые одноклеточный организм «принимает» под воздействием окружающей среды, – быстрые, химические и внутренние по отношению к отдельной клетке. Высшие организмы и одноклеточные добывают информацию из внешнего мира очень похожими способами, но вот обрабатывают они ее по-разному.
Многоклеточная жизнь появилась почти на 1,5 миллиарда лет позже одноклеточной. Сегодня существует большое количество одноклеточных эукариот, и схема их родства ясно показывает, что уже на ранних этапах эволюции было много случаев дивергенции между ними и многоклеточными животными и растениями. Это наблюдение справедливо, потому что не все одноклеточные эукариоты – ближайшие родственники и не все многоклеточные организмы происходят от одного общего предка. Некоторые одноклеточные эукариоты, например, более тесно связаны с растениями, чем с другими одноклеточными эукариотами. Взять хотя бы водоросли и одноклеточные эукариоты – хламидомонады («хлами», как ласково называют их ученые): и те и другие гораздо теснее связаны с растениями, чем с другими одноклеточными организмами, такими как амеба.
Растения довольно хорошо понимают сигналы раздражителей из окружающего мира, но для этого они развили иные механизмы, нежели животные. Отличный пример – подсолнух: если можете, потратьте несколько часов, наблюдая, как он реагирует на солнечный свет. Самый интересный момент происходит на рассвете, когда цветок медленно поворачивается туда, где ожидается восход солнца. Подсолнух очень точно определяет время и ловко передвигает свой цветочек на нужное место. Другой пример – мимоза, растение, реагирующее на прикосновения. А любой, кто видел «Лавку ужасов», легко вспомнит венерину мухоловку, которая быстро и жадно хватает добычу, необдуманно оказавшуюся поблизости с ее ловушкой (рис. 1.2). Однако у растений нет нервных клеток и, следовательно, нет мозга или нервной системы, как у животных. (Я делаю эти категоричные заявления о растениях и нервных системах, хотя знаю, что существует журнал под названием Plant Neurobiology («Нейробиология растений») и несколько институтов занимаются изучением одноименной науки. Но нейробиология растений и нейробиология животных все же фокусируются на разных факторах.)
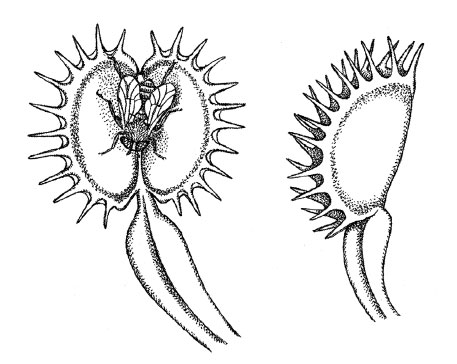
Рис. 1.2. Нейробиология растений или интеллектуальное поведение? Венерина мухоловка (Dionaea muscipula)
Метафора становится важной при изучении реакции организмов на окружающую среду. Организм с «метафорическим мозгом», подобный растению, не обрабатывает информацию из внешнего мира так, как это делают позвоночные, и это неудивительно. Под метафорическим мозгом я подразумеваю систему, аналогичную мозгу позвоночных, но не являющуюся нейронной. Именно эта способность реагировать на внешний мир побудила некоторых исследователей инициировать нейробиологический подход по отношению к растениям. Но очень трудно отрицать, что у растений нет мозга и нервной системы. Я предпочитаю признать, что растения довольно хорошо чувствуют внешний мир и у них есть некий способ интегрировать свое восприятие окружающей среды, но в функциональном структурном контексте у них нет мозга. С точки зрения эволюционной биологии можно сказать, что растительная версия нервной системы приближена к мозгу насекомого или позвоночного. Центральная сенсорная система растения – это метафора нервной системы беспозвоночных или позвоночных. С интеллектуальной точки зрения мне гораздо приятнее осознавать, что растения изобрели новый способ восприятия внешнего мира, не имеющий ничего общего с нервной системой. И действительно, когда мы начинаем исследовать пути, по которым развивались структуры и механизмы традиционных чувств у животных с нервной системой, эта гипотеза многократно подтверждается. Вероятно, надо согласиться с Майклом Полланом, ярым защитником растительной жизни на планете, который предлагает говорить не о «нейробиологии растений», а скорее о «разумном поведении растений». И в этом контексте растения развили свои способности особым путем, без какого-либо эволюционного сходства с разумным поведением животных, за исключением использования некоторых основных молекулярных инструментов эволюционного инструментария, который есть у большинства многоклеточных эукариот. Разумное поведение позволяет растению воспринимать стимулы из окружающей среды, такие как свет или концентрация химических соединений, и интерпретировать их «интеллектуальным» образом. Нейронная основа разумного поведения растений – это просто еще одно решение для межклеточной коммуникации, на которое натолкнулась жизнь на Земле и которое развивалось как ответ на потребность в сенсорной связи с внешним миром.
Неудивительно, что организмы, не имеющие глаз, ушей, носа, кожи и рта, не могут видеть, слышать, обонять, осязать и ощущать вкус. Эти так называемые традиционные, или аристотелевские, чувства – сфера компетенции развитых животных. И потому организмы без этих функций фокусируют свои ощущения на других раздражителях окружающей среды, таких как электрические и магнитные поля и химические сигналы, которые ведут себя не так, как вкус и запах. Организмы, которые могут видеть, слышать, обонять, осязать и ощущать вкус, развили удивительный набор анатомических и физиологических особенностей, которые усиливают эти чувства. И разнообразие стимулов окружающей среды, которые жизнь подарила нам для восприятия внешнего мира, поражает воображение.
2. Мозг и первомозг
Мозг (или его отсутствие) – от губки до человека
Губка видит все? Губка ничего не видит.
Лоуренс Тирни, актер
Древо жизни росло и разветвлялось, а на пути развития чувств возникали определенные этапы. К самым ранним из них относится эволюция межклеточной коммуникации посредством молекулярных процессов, таких как чувство кворума. Затем пришло время наборов генов, отвечающих за выполнение специфических клеточных функций, то есть того самого молекулярного инструментария, благодаря которому информация передается через одиночные клетки. Потом эволюционировали многоклеточные организмы, клетки которых стали взаимодействовать между собой с помощью сигнальной трансдукции. После возникновения многоклеточности открылись новые возможности коммуникации клеток, и они значительно повлияли на биологию наших прямых предков-эукариот. К важным вехам эволюции также относятся потенциал действия, синапс, дифференцированные нервные клетки, нейронные сети, скопления нейронов и специализированная нервная система. И, несмотря на то что все эти этапы развития (то есть все, чего мы достигли в процессе эволюции нашей ветви) могут показаться дорогой к совершенству, это всего лишь промежуточные остановки. Другие ветви тоже развивались, но стадии эволюции нашей нервной системы не имеют к ним ни малейшего отношения.
Чарльз Дарвин уподобил отношения организмов с «великим древом жизни», чтобы наглядно показать взаимосвязь всех организмов на планете через общее происхождение и дивергенцию. Биологи взяли на вооружение концепцию «великого древа жизни» для отслеживания эволюции признаков. Наблюдая за случаями дивергенции организмов и признаков, имеющих отношение к нервной системе, мы получаем довольно точную картину, как развивался мозг в целом и наш мозг в частности.
Самые ранние ветви древа, иллюстрирующего эволюцию животных, включают губку и небольшие организмы, относящиеся к типу пластинчатых и по форме напоминающие блин. Некоторые исследователи полагают, что все губки – родственники друг другу благодаря общему предку. Другие же считают, что существовали две линии развития губок. Доподлинно известно, что губки имеют примерно восемь типов клеток и ни один из них не является нейронным, именно поэтому у них и нет мозга. У пластинчатых, в свою очередь, четыре типа клеток, среди которых нет даже одного нейронного, поэтому и у них тоже не может быть мозга. Но вот что интересно: у этих двух древних животных, находящихся у основания древа, есть большое количество генов, необходимых для создания нервных клеток, просто они их не используют. По всей видимости, они нашли другое применение генам, которые другие животные направляют на строительство нейронов. И губки, и пластинчатые чувствуют изменения окружающей среды и реагируют на них, поэтому эти организмы можно назвать чувствующими, несмотря на отсутствие мозга и нервной системы. Губки «чихают», когда ощущают контакт с какими-то частицами. Это «чихание» представляет собой очень эффектное зрелище, но весь процесс занимает около часа, поэтому наблюдать за губками лучше через замедленную съемку. Пластинчатые очень охочи до еды и весьма преуспели в ее добыче – и это при отсутствии мозга.
Гребневики, или ктенофоры, – невероятно интересные существа. Выглядят они чертовски круто, и считается, что это первая группа животных, отклонившаяся в развитии от одноклеточных предков – эукариот. У них есть несколько типов клеток, в том числе и нервные клетки, есть нервная система, но нет централизованного скопления клеток, которые формируют мозг. Если ктенофоры действительно являются первыми животными, чья эволюция устремилась по другой ветке древа жизни, то тогда вырисовывается любопытный сценарий развития событий. Первый момент: нервные системы гребневиков и других животных очень схожи (за исключением губок и пластинчатых). Второй момент: нервную систему относительно легко утратить, как это случилось в ветви, по которой развивались губчатые и пластинчатые. Этот пример показывает, как применение концепции общих предков и древа жизни позволяет нам проанализировать значимые изменения нервной системы, произошедшие при дивергенции животных.
2.1 Потенциал действия
Потенциал действия – это физиологическая реакция клетки, при которой электрический разряд в ней повышается и понижается. У более сложных животных есть регулируемая мембраной разница электрического потенциала между внутренней и внешней сторонами клетки. Мембрана поддерживает напряжение внутри клетки на уровне минус 70 милливольт относительно внешней среды, и это называется состоянием покоя. Когда клетку каким-то образом стимулируют извне, разница потенциалов снаружи и внутри повышается примерно на 110 милливольт и становится равна 40 милливольтам. В результате заряд внутри клетки повышается. Затем напряжение резко падает гораздо ниже значения состояния покоя, а после возвращается к изначальным 70 милливольтам. Повышение и понижение потенциала действия происходит очень последовательно, и именно так нервные импульсы или электрические сигналы передаются от клетки к клетке по всей нервной системе. Потенциалы действия также важны для связи нервной системы с сенсорной и моторной системами.
Однако способы взаимодействия нервных клеток у нашей ветки и более низших, «мягких», животных одни и те же. Похоже, что передача электрического сигнала посредством потенциала действия – это изобретение древних животных.
После того как отделились ветви гребневиков, губчатых и пластинчатых, общий предок стрекающих кишечнополостных и группа билатеральных, к которой относимся и мы, тоже разошлись. Любой организм, симметричный относительно линии, проведенной от центра головы до конца тела, называется билатеральным или двусторонне-симметричным. Наиболее близкородственная билатеральным группа – это стрекающие кишечнополостные, книдарии, объединяющие медуз, гидр, кораллы и очень странную группу организмов, называющихся кубомедузами из-за их формы, напоминающей коробку. У стрекающих кишечнополостных есть клетки, которые можно назвать нервными, и нервная система, которую лучше всего описывать словом «сеть», но нет централизованного сосредоточения нервных клеток, которое можно называть мозгом.
Билатеральные делятся на две большие группы: первичноротые (например, насекомые и моллюски) и вторичноротые (куда входит человек). В организмах обеих основных групп билатеральных есть мозг, поэтому ученые обычно считают его признаком именно этого таксона[5]. Большинство биологов склонны описывать мозг первично- и вторичноротых, используя термин «гомологичный». Дарвин был первым, кто придал ему смысл, указав, что гомологичные признаки – это те, что передаются в результате прямого наследования от общего предка. К примеру, крылья птицы и летучей мыши не гомологичны, поскольку существует множество бескрылых млекопитающих, прерывающих общую родословную этих двух видов позвоночных. Любые признаки, присутствующие у организмов, но не связанные одной линией происхождения, считаются аналогичными и возникли в результате конвергенции (сходства).
Но действительно ли мозг можно назвать признаком билатеральных? Считается, что общая структура мозга первичноротых отличается от более развитого мозга вторичноротых. У общего предка билатеральных была нервная сеть, которая охватывала большую часть его организма, а в ее переднем отделе находилось некое скопление нервных клеток, которое нейробиологи назвали бы мозгом. Некоторые нейроанатомы – специалисты по нервной системе – предполагают, что общий предок билатеральных даже имел сложную структуру, состоявшую из трех отделов.
Вполне возможно, что скопление клеток в переднем отделе нервной системы предка билатеральных начало развиваться в двух направлениях, что привело к появлению первичноротых и вторичноротых. Чтобы как-то отличать мозг первичноротых от развитого и структурированного мозга вторичноротых, я часто называю тот первомозгом. Мы можем использовать насекомых в качестве типовой системы для расшифровки того, как сенсорная информация собирается из внешнего мира и превращается в нервные импульсы, но, как только та попадает в мозг в случае вторичноротых или в первомозг у первичноротых, сходство между этими двумя типами нейронных центров заканчивается. Вероятнее всего, сбор информации органами чувств и передача ее для обработки – это единственное, что можно сопоставить у мозга и первомозга. Судя по всему, наши билатеральные предки обладали довольно обширным арсеналом сенсорных инструментов, и наблюдения за такой удивительной дифференциацией мозга и первомозга после дивергенции билатеральных ясно показывают, что этот инструментарий использовался различным образом по мере того, как один вид появлялся вслед за другим. Справедливости ради надо отметить, что некоторые исследователи приводят доводы в пользу гомологии на основе другой важной информации. Первичноротые, такие как фруктовая мушка дрозофила (Drosophila melanogaster), для нейронной обработки используют многие из тех же генов, что и вторичноротые. Кроме того, они имеют очень похожие, хотя и неидентичные структуры, где нервные клетки вступают в контакт друг с другом (так называемые синапсы – см. вставку 2.2). Но, если вдуматься, это любопытная мысль: сложный мозг сформировался как минимум дважды на этой планете – в ветке первичноротых и у вторичноротых (рис. 2.1).
2.2 Синапсы
Синапсы – это точки контакта, которые позволяют одному нейрону связываться с другим посредством передачи электрического сигнала. Механизм появления этого сигнала довольно сложный (см. вставку 2.1). Клетка, передающая электрический заряд, называется пресинаптической, а получающая его – постсинаптической. Ученые полагают, что взаимодействие клеток других типов, таких как мускульные клетки, с нейронами тоже осуществляется через синапсы.
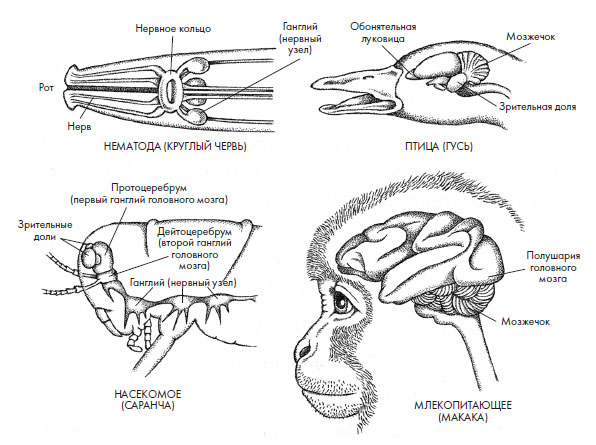
Рис. 2.1. Сравнение мозга позвоночных (птицы и примата) с мозгом нематоды (круглого червя) и насекомого
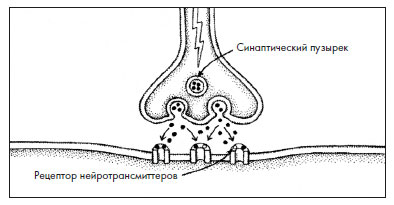
Рис. 2.2. Структура синапса. Пресинаптическая клетка содержит синаптический пузырек (везикулу), выпускающий нейротрансмиттеры, а мембрана постсинаптической клетки содержит клеточный рецептор – белок
Если не считать мозг первичноротых тем же типом мозга, что и наш, или по крайней мере не называть его триединым, то можно использовать мозг, состоящий из трех частей, в качестве эвристического приема, чтобы понять, как эволюционировал мозг позвоночных. У организмов ранних веток вторичноротых, таких как морские звезды или морские ежи, есть нервные клетки, но они распределены по всему телу без центрального скопления клеток, которое можно было бы назвать мозгом. Некоторые ученые проявляют верх педантизма и считают, что у этих организмов, известных также как иглокожие, есть своего рода «распределенный мозг».
Следующей группой животных, которые поучаствовали в «подвешивании» человеческого мозга на древо жизни, считаются родственники иглокожих – хордовые (Chordata). У всех ее представителей, самыми первыми из которых были личиночнохордовые и головохордовые, есть нервные трубки. Личиночнохордовые, или оболочники, – довольно странные существа. Ярким представителем этого подтипа можно назвать одиночную асцидию, относящуюся к группе туникатов, получивших это название из-за защитной оболочки (туники), которая образуется у взрослых особей из материала, похожего на целлюлозу. Одиночная асцидия (как и земноводные) проходит личиночную стадию, во время которой развивается мозговой пузырь, и это позволяет нам говорить, что у нее уже есть место для мозга, хотя самого мозга там нет. Как ни странно, но, превращаясь во взрослую особь, личинка одиночной асцидии устраивает пир из собственного мозга. У личинки нет внешнего рта, чтобы заглатывать пищу, и поэтому она вынуждена искать иной способ, чтобы ее получить. А поскольку в дальнейшем крошечный комочек нервных клеток, составляющих мозговой пузырь, никак не используется взрослой асцидией, личинка реабсорбирует эту ткань. Да, мозга у них нет, но зато есть нотохорд, хотя это еще и не позвоночный столб, который на древе жизни появляется позже. У головохордовых тоже есть нервные трубки, но опять-таки нет позвоночника. Трудно однозначно утверждать, что у одного из лучших представителей этого подтипа – ланцетника – есть мозг, и на практике анатомы называют то, что можно у него считать таковым, утолщением нервной трубки. Ведь в буквальном смысле это просто небольшая выпуклость на нотохорде в области головы ланцетника.
У позвоночных есть позвоночный столб, или позвоночник, и полностью развитая нервная трубка. Одно время нейробиологам было трудно смириться с тем, что эволюция мозга позвоночных была линейной с поэтапным добавлением все более сложных деталей. Лимбическая система и кора головного мозга – два наглядных примера более сложно организованных нейронных образований, которые постепенно появились у общих предков в результате дивергенции. Можно было бы предположить, что усложненные структуры или процессы должны развиваться из простых, но на практике совсем не обязательно, что потомки всегда устроены сложнее, чем их предки. Да и результатом эволюции может быть некая упрощенная версия прототипа. Тем не менее, если отправной точкой становится примитивная структура, единственный путь ее развития – обрастать дополнительными деталями. И кажется, мозг позвоночных эволюционировал именно так: путем наслоения новых структур. Проблема в том, что непонятно, как и когда были добавлены эти слои. Схемы эволюции мозга всегда включают в себя изображение трех его основных частей, что приводит к теории триединого мозга. По определенным критериям выделяют три главных отдела мозга: ствол и мозжечок (называемые Р-комплексом, где «Р» – от слова «рептилия»), лимбическую систему и кору. Это довольно произвольное деление, и, видимо, именно поэтому некоторые нейробиологи отрицают трехчастное строение мозга. Тем не менее в качестве эвристического подхода использование триединой модели мозга позвоночных работает довольно неплохо. Но даже с учетом этого все равно остается спорным вопрос: как развивался трехчастный мозг и действительно ли он трехчастный?
Мозг предков всех позвоночных выглядел довольно примитивно, и у него либо не было коры, либо она была очень маленькая, а мозжечок находился в зачаточном состоянии. В основном поведение предков контролировалось стволом мозга и мозжечком, отвечающими за основные функции организма, такие как сердцебиение и дыхание. Обладали предки и базовыми способностями реагировать на внешний мир: мозг мог распознавать запахи, звуки и объекты. Это было то, что в классической теории триединого мозга называется рептильным мозгом. Последними слоями в схеме трехчастного мозга стали мозги палеомлекопитающих и неомлекопитающих. Большинство из тех, кто работает в сфере биологии развития, исходя из этих названий сделает вывод, что два следующих важных предка – это давно вымершие млекопитающие (млекопитающие палеоцена и неоцена). Недостаток этой схемы в том, что в ней отсутствуют птицы, у которых довольно сложные нейронные способы взаимодействия с внешним миром и чей мозг не вписывается в общий континуум. Поэтому необходимо либо пересмотреть названия предков мозга, либо предположить, как мы уже делали раньше, что мозг птиц – это другая надстройка над мозгом ящериц и его не следует отождествлять с мозгом млекопитающих. Можно переименовать рептильный мозг в мозг рыб и попытаться сделать шаг в сторону на древе жизни. Вполне возможно, что следующим будет рептильный/птичий мозг, а затем – прото/неомозг млекопитающих. Проблема этой теории в том, что некоторые биологи-эволюционисты не выделяют рыб в отдельную группу. Почему? Несмотря на то что название «рыба» носит дескриптивный характер, оно не дефинитивно, то есть не определяет какую-то группу организмов, отличающуюся от других по неким признакам, которые обычно учитывают ученые-таксономисты при систематизации. Но решить эту проблему довольно просто: надо или окрестить основные рода рыб как-то иначе, или называть всех потомков общего предка тех, кого мы привыкли считать рыбой, рыбами. Включая людей, разумеется. Так выплюнуть наживку или заглотить ее? Пока давайте-ка не будем спешить попасться на крючок и посмотрим на триединый мозг в ином ракурсе.
Если бы кто-то проанализировал все названия, которые нейробиологи давали трем главным отделам мозга, он столкнулся бы как с неким произволом, так и с высокой степенью субъективности – в зависимости от того, какие именно виды имеют те или иные структуры. И это недвусмысленно свидетельствует, что многим ученым нравится идея существования трех основных частей мозга (рис. 2.3).
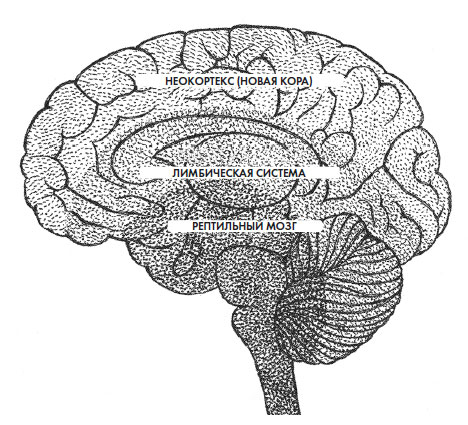
Рис. 2.3. Триединый мозг
Рассмотрим дивергенцию позвоночных в надежде, что это может придать нашему обсуждению некоторую объективность. После того как у общего предка рыб, амфибий, рептилий, птиц и млекопитающих сформировались основные анатомические структуры основы головного мозга (внутренний мозг рыбы), появился следующий общий предок, породивший мозг всех высших позвоночных (рептилий, птиц и млекопитающих). И на этом этапе добавился еще один слой мозга, позволивший использовать более сложные способы обработки сигналов внешнего мира, поступающих от органов чувств, – наш рептильный мозг. Это привело к развитию внутренней области мозга, которая включала в себя то, что обычно называют лимбической системой. Подобный порядок отлично вписывается в теорию триединого мозга, если учесть, что лимбическая система есть у всех вышеупомянутых позвоночных. Впоследствии лимбическая система стала решать очень интересные задачи, связанные с реагированием на окружающую среду. Последней сформировалась кора головного мозга, которая у всех потомков общего предка разрасталась по-разному. Важно отметить, что неокортекс специфическим образом изменяется у потомков того самого общего предка, о чем свидетельствуют экспансия неокортекса у птиц и млекопитающих и отсутствие его расширения у рептилий. Короче говоря, неокортекс птиц – это другой вид расширения коры, отличный от экспансии неокортекса млекопитающих.