Р. П. Самусев
Общая и частная гистология
2.1. Клеточный цикл
Клеточный цикл (КЦ; cyclus cellularis) – совокупность явлений между двумя последовательными делениями клетки или между ее образованием и гибелью (рис. 2.5).
В ходе КЦ обеспечивается функция воспроизведения клеток и передачи генетической информации. КЦ включает собственно митотическое деление и интерфазу – промежуток между делениями.
Интерфаза включает пресинтетический, или постмитотический (G1), синтетический (S) и постсинтетический, или премитотический (G2), периоды. В интерфазе клетка увеличивается в размерах и удваивает генетический материал.
В большинстве тканей делится лишь небольшая часть клеток, остальные дифференцируются и пребывают в G0-периоде.
G1-период – промежуток сразу после митотического деления клетки; характеризуется активным ростом клетки и синтезом белка и РНК, благодаря чему дочерние клетки достигают нормальных размеров и восстанавливают необходимый набор органелл. В этот период синтезируются особые «запускающие белки», или активаторы S-периода, которые обеспечивают переход клетки в S-период.
S-период характеризуется удвоением (репликацией) ДНК и синтезом белков (гистонов), обеспечивающих нуклеосомную упаковку вновь синтезированной ДНК. Одновременно удваивается число центриолей. S-период у большинства клеток длится 8—12 ч.
G2-период продолжается вплоть до митоза. В течение этого периода клетка готовится к делению: происходит созревание центриолей, запасается энергия, синтезируются РНК и белки (тубулины). Длительность G2-периода составляет 2–4 ч.
За G2-периодом следует митоз. Он завершает КЦ, образуется две идентичные (дочерние) клетки.
Митоз (mitosis; кариокинез, или непрямое деление клетки) является универсальным механизмом деления клеток. Он включает основные фазы: профазу, метафазу, анафазу и тело-фазу (см. рис. 2.5).
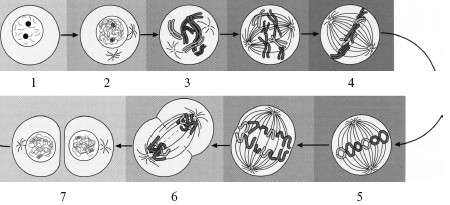
Рис. 2.5. Клеточный цикл (схема).
1 – интерфаза; 2 – профаза; 3 – прометафаза; 4 – метафаза; 5 – анафаза; 6 – телофаза; 7 – цитокинез.
Профаза начинается с конденсации хромосом, которые под световым микроскопом предстают в виде нитевидных структур.
Каждая хромосома состоит из двух параллельно лежащих хроматид, связанных друг с другом с помощью суженного участка – центромеры.
К концу профазы ядрышко и ядерная оболочка исчезают, а центриоли мигрируют к противоположным полюсам клетки и дают начало нитям митотического (ахроматинового) веретена. В области центромеры образуются особые белковые комплексы – кинетохоры, которые прикрепляют хроматиды к нитям веретена.
Метафаза соответствует максимальной конденсации хромосом. Они выстраиваются в области экватора митотического веретена в виде экваториальной (метафазной) пластинки (вид сбоку) или материнской звезды (вид со стороны полюсов), удерживаемые здесь благодаря сбалансированному натяжению кинетохорных микротрубочек.
Сестринские хроматиды в конце этой фазы разделяются щелью, соединенные только в области центромеры.
Анафаза начинается с синхронного расщепления всех хромосом на сестринские хроматиды (в области центромера) и движения дочерних хромосом к противоположным полюсам клетки. Характеризуется удлинением митотического веретена за счет некоторого расхождения полюсов клетки. Завершается скоплением на полюсах клетки двух идентичных наборов хромосом (стадия дочерних звезд).
В конце анафазы благодаря сокращению актиновых микро-филаментов, концентрирующихся по окружности клетки (сократимое кольцо), начинает образовываться клеточная перетяжка.
Телофаза – конечная стадия митоза, в течение которой реконструируются ядра дочерних клеток и завершается их разделение. Вокруг хромосом восстанавливается кариолемма, с которой связывается формирующаяся ядерная пластинка, вновь появляются ядрышки. Ядра дочерних клеток постепенно увеличиваются, а хромосомы прогрессивно деспирализуются и исчезают, замещаясь картиной хроматина интерфазного ядра. Клеточная перетяжка углубляется, так что дочерние клетки в течение некоторого времени остаются связанными только узким мостиком из пучка микротрубочек – срединным тельцем; дальнейшая перешнуровка цитоплазмы завершается образованием двух дочерних клеток.
В телофазе происходит также распределение органелл между дочерними клетками (митохондрий, ЭПС, комплекс Гольджи).
Эндомитоз – вариант митоза, при котором происходит удвоение числа хромосом внутри ядра без разрушения кариолеммы и образования веретена деления, что приводит к значительному увеличению содержания ДНК в ядре – полиплоидии и увеличению его объема.
Наличие полиплоидных клеток – нормальное явление в печени, эпителии мочевого пузыря, клеток концевых отделов слюнных желез, поджелудочной железы.
Основной смысл развития полиплоидии заключается в усилении функциональной активности клетки.
Общий контроль активности деления клеток обеспечивают протоонкогены, антионкогены, факторы роста (факторы роста нервов, эпидермальный фактор роста, тромбоцитарный фактор роста, инсулиноподобные факторы роста, колониестимулирующие факторы и др.), а также кейлоны – гормоноподобные регуляторы, угнетающие клеточное размножение.
2.2. Старение и гибель клеток
После функционирования в течение определенного периода времени клетка стареет и гибнет.
Морфологическими признаками старения клетки являются уменьшение ее объема, редукция большинства органелл, увеличение содержания лизосом, накопление пигментных и жировых включений, нарастание проницаемости клеточных мембран, вакуолизация цитоплазмы и ядра.
Гибель клеток обеспечивается двумя видами морфологических изменений, которые соответствуют различным механизмам ее развития – некрозом и апоптозом.
Некроз возникает под действием выраженных повреждающих факторов (перегревание, переохлаждение, недостаток кислорода, нарушение кровоснабжения, механические травмы и т. п.).
При некрозе происходит разрушение клеточных структур после выделения гидролаз и других ферментов из поврежденных лизосом, кариопикноз, кариорексис и кариолизис ядра, исчезновение клеточных границ и распад клетки.
Апоптоз – физиологическая (запрограммированная) гибель клеток. Это активный энергоемкий генетически контролируемый процесс, регулируемый внутренней программой, которая запускается внешними факторами.
При апоптозе клетка теряет все специализированные структуры на своей поверхности (микроворсинки и межклеточные соединения), происходит уплотнение цитоплазмы и ядра. Конденсация цитоплазмы приводит ко все более компактному расположению органелл, которые в отличие от некроза сохраняют свою целостность.
Изменения в ядре включают только кариопикноз и кариорексис (без разрушения кариолеммы), кариолизис отсутствует; хроматин в ядре укладывается в виде крупных полулуний, после чего ядро распадается на фрагменты.
Плазмолемма клетки образует многочисленные вздутия и выпячивания, содержащие органеллы и фрагменты ядра, которые отшнуровываются, формируя округлые или овальные апоптозные тела. Последние захватываются соседними клетками посредством фагоцитоза и перевариваются ими.
Апоптоз – один из фундаментальных и универсальных механизмов тканевого гомеостаза, который наблюдается в различных тканях человека и животных в норме, патологии, эмбриональном развитии и у взрослого.
Тесты и вопросы для самоконтроля
Выберите правильные ответы.
1. Плазматическая мембрана:
а) связана с определенными компонентами цитоскелета;
б) состоит из трех липидных слоев;
в) содержит на наружной поверхности гликокаликс;
г) не дает возможности белкам перемещаться в толще мембраны.
2. Ядерная пора:
а) шестиугольная;
б) ограничена одинарной мембраной;
в) является структурой скорее изменчивой, нежели стабильной;
г) делает возможным сообщение между ядром и цитоплазмой.
3. Рибосомы:
а) прикреплены к поверхности внутренней ядерной мембраны;
б) организованы в полисомы в клетках, синтезирующих белок для собственных нужд;
в) всегда связаны с иРНК;
г) состоят из большой и малой субъединиц.
4. Агранулярная (гладкая) эндоплазматическая сеть:
а) часто выглядит как разветвляющиеся и анастомозирующие друг с другом трубочки;
б) иногда несет на своих мембранах рибосомы;
в) присутствует в клетках, где происходит детоксикация лекарственных средств;
г) редко наблюдается в клетках скелетных мышц.
5. Комплекс Гольджи:
а) имеет конденсированные вакуоли, связанные с его наружной (цис-) поверхностью;
б) имеет конденсированные вакуоли, связанные с его внутренней (транс-) поверхностью;
в) синтезирует мультивезикулярные тельца;
г) участвует в синтезе некоторых липопротеинов.
6. Внутриклеточное переваривание:
а) связано с лизосомами;
б) включает процесс аутофагии;
в) происходит в комплексе Гольджи;
г) участвует в обновлении состава органелл, их переработке.
7. Цитоскелет:
а) включает микротрубочки;
б) включает гликокаликс;
в) включает промежуточные филаменты;
г) включает актиновые филаменты.
8. Ядерная оболочка:
а) отсутствует у всех эпителиальных клеток;
б) содержит эухроматин;
в) иногда продолжается в цистерны комплекса Гольджи;
г) сформирована двумя ядерными мембранами.
9. Скорее включениями, чем органеллами, являются все перечисленные структуры, кроме:
а) капли жира;
б) лизосома;
в) гликоген;
г) кристаллоид;
д) гранула слизи.
10. Ядрышко характеризуется всеми перечисленными структурами, кроме:
а) фибриллярный компонент, представляющий собой ранние стадии формирования предшественников рРНК;
б) хроматин, связанный с ядрышком;
в) ядерная пластинка;
г) гранулярный компонент, представляющий собой поздние стадии формирования предшественников рРНК.
11. Из всех перечисленных утверждений о лизосомах неверно то, что они:
а) связаны с активностью кислой фосфатазы;
б) участвуют во внутриклеточном пищеварении;
в) содержат ряд гидролитических ферментов;
г) могут быть идентифицированы по цитохимической реакции на каталазу;
д) часто выглядят как плотные, окруженные мембраной тельца.
12. Митоз в клеточном цикле не включает следующей фазы:
а) анафаза;
б) профаза;
в) метафаза;
г) интерфаза;
д) телофаза.
13. Цитохимическое исследование выявило в цитоплазме клетки большое содержание гидролитических ферментов. Об активности каких органелл свидетельствует этот факт?
14. Известно, что в живой клетке происходит постоянное перемещение органелл. Какие структурные элементы клетки принимают в этом участие?
15. В клетку проник фактор, нарушающий целостность мембран лизосом. Какие изменения произойдут в клетке?
16. В процессе жизнедеятельности клетки резко увеличивается число цистерн и канальцев агранулярной эндоплазматической сети. Синтез каких веществ активизируется в клетке?
17. На клетки подействовали препаратом, изменяющим структуру рибосом. Какие процессы будут нарушены в первую очередь?
18. С помощью микроманипулятора из клетки удалили комплекс Гольджи. Как это отразится на ее дальнейшей жизнедеятельности?
19. Клетку обработали препаратом, блокирующим функцию ядрышка. Как это отразится на жизнедеятельности клетки?
20. На препарате видна митотически делящаяся клетка (диплоидная) на стадии анафазы. Сколько хромосом входит в состав каждой дочерней звезды?
Ответы
1: а, в.
2: г.
3: б, в, г.
4: а, в.
5: б, г.
6: а, б, г.
7: а, в, г.
8: г.
9: б.
10: в.
11: г.
12: г.
13. Об активности лизосом.
14. Микрофиламенты и микротрубочки.
15. Аутолитические. Аутолиз.
16. Липидов и углеводов.
17. Синтез белков (процессы трансляции).
18. Нарушатся синтез углеводов, образование лизосом, упаковка, созревание и выведение секреторных продуктов клетки.
19. Нарушатся образование рибосом и синтез белков.
20. 46 хромосом.
Глава 3
Эпителиальные ткани
Эпителиальные ткани (ЭТ; textus epitheliales) – это ткани, выстилающие внутренние органы и полости тела и покрывающие наружную его поверхность, а также образующие большинство желез. Характеристика ЭТ:
♦ специализированы для выполнения различных функций: абсорбция, секреция, экскреция, транспортная, сенсорная, защитная и т. д.;
♦ состоят из специализированный; клеток – эпителиоцитов, лежащих в один слой (однослойный эпителий) или в несколько слоев (многослойный эпителий), а также рядов (многорядный эпителий);
♦ клетки расположены тесно друг к другу с узкими межклеточными промежутками между ними;
♦ не содержат сосудов, но обладают высокой способностью к регенерации;
♦ эпителиоциты характеризуются полярностью, наличием развитых межклеточных соединений и специализированы для выполнения разнообразных функций переноса;
♦ отделены от подлежащей рыхлой соединительной ткани особым структурным слоем – базальной мембраной (пластинкой).
Выделяют поверхностный (покровный), железистый, чувствительный и герминативный эпителий (схема 3.1).
3.1. Поверхностный эпителий (epithelium superficiale)
Однослойный плоский эпителий (рис. 3.1):

Рис. 3.1. Однослойный плоский эпителий (мезотелий сальника); тотальный препарат. ×300.
1 – эпителиоцит; 2 – ядро; 3 – клеточные границы.
• состоит из одного слоя плоских клеток;
• выстилает кровеносные сосуды (эндотелий), плевральную, брюшинную и другие серозные полости (мезотелий);
• образует париетальный слой почечной капсулы Боумена – Шумлянского, петли Генле нефрона и т. д.
Схема 3.1. Виды эпителия


Однослойный кубический эпителий (рис. 3.2):

Рис. 3.2. Однослойный кубический эпителий канальцев почки. ×300.
1 – просвет канальца; 2 – базальная мембрана; 3 – эпителиальные клетки; 4 – ядро.
• состоит из одного слоя многогранных клеток, которые на гистологических срезах выглядят кубическими;
• выстилает дистальные части почечных канальцев, фолликулы щитовидной железы, поверхность яичника и т. д.
Однослойный призматический эпителий (рис. 3.3):

Рис. 3.3. Однослойный призматический эпителий канальцев почки. ×500.
1 – просвет канальца; 2 – базальная мембрана; 3 – апикальный полюс эпителиоцита; 4 – ядро эпителиоцита; 5 – базальный полюс эпителиоцита.
• состоит из многогранных клеток, вытянутых в одном направлении и имеющих на гистологических срезах вид призм или цилиндров;
• клетки расположены в один слой;
• выстилает желудок, тонкую и толстую кишки и экскреторные протоки многих желез.
Выделяют призматический реснитчатый (эпителий матки, маточной трубы) и безреснитчатый (эпителий желудка, тонкой кишки) виды.
Многорядный эпителий (рис. 3.4):

Рис. 3.4. Однослойный многорядный реснитчатый эпителий трахеи. ×440.
1 – эпителиоциты; 2 – реснички; 3 – базальная мембрана.
• является одним из видов однослойного эпителия, так как каждая клетка лежит на базальной мембране, но не все из них достигают просвета полого органа;
• выстилает трахею, крупные бронхи, выделительные протоки околоушной железы и т. д.
Различают многорядный реснитчатый (эпителий трахеи, крупных бронхов) и безреснитчатый (эпителий междольковых протоков околоушной железы) виды.
Многослойный плоский неороговевающий эпителий (рис. 3.5):

Рис. 3.5. Многослойный плоский эпителий роговицы глаза (передний эпителий). ×270.
1 – базальная мембрана; 2 – базальные клетки; 3 – шиповатые клетки; 4 – кроющие клетки.
• состоит из нескольких слоев клеток различной формы;
• самый поверхностный слой состоит из плоских ядерных клеток;
• выстилает влажные поверхности тела (полость рта, пищевод, влагалище, роговицу).
Многослойный плоский ороговевающий эпителий (рис. 3.6):

Рис. 3.6. Многослойный плоский ороговевающий эпителий кожи пальца (эпидермис).×250.
1 – базальная мембрана; 2 – базальный слой; 3 – шиповатый слой; 4 – зернистый слой; 5 – блестящий слой; 6 – роговой слой.
• состоит из нескольких слоев разнообразных клеток;
• уплощенные клетки поверхностного слоя не имеют ядер и заполнены кератином (т. е. ороговели);
• образует эпидермис, выстилающий наружную поверхность тела.
Многослойный кубический эпителий:
• состоит из двух и более слоев клеток. Клетки поверхностного слоя выглядят кубическими;
• выстилает протоки большинства потовых желез кожи.
Многослойный призматический эпителий:
• состоит из двух или более слоев клеток. Клетки поверхностного слоя в большинстве своем выглядят призматическими;
• встречается в крупных выводных протоках некоторых желез и в пещеристой части мужского мочеиспускательного канала.
Переходный эпителий (рис. 3.7):

Рис. 3.7. Переходный эпителий мочевого пузыря (в спавшемся состоянии). ×420.
1 – базальная мембрана; 2 – базальный слой; 3 – промежуточный слой; 4 – поверхностный слой.
• многослойный эпителий, клетки которого в поверхностном слое в расслабленном состоянии имеют форму купола и выдаются в просвет полости органа;
• в растянутом состоянии представляет собой несколько слоев плоских клеток;
• претерпевает отмеченные выше изменения в зависимости от степени наполнения органа (например, мочевого пузыря);
• выстилает мочевыводящие пути (от почечных чашек до уретры).
Эпителиальные клетки соединяются друг с другом с помощью различных типов (механических и коммуникационных) мембранных соединений. Различают соединения:
• простое межклеточное (junctio intercellularis simplex), при котором плазмолеммы соседних клеток сближены на расстояние 15–20 нм. Выделяют:
а) зубчатое соединение (junctio intercellularis denticulata);
б) пальцевидное соединение (junctio intercellularis digitiformis);
• сложное межклеточное, или соединительный комплекс (junctio intercellularis complex);
• десмосому (desmosoma);
• нексус, или щелевидное соединение (nexus; macula communicans).
Сложное межклеточное соединение, или соединительный комплекс, – это специализированный участок между боковыми поверхностями эпителиальных клеток, который является барьером между внешней и внутренней средой (например, в тонкой кишке).
На электронных микрофотографиях четко выделяются 3 участка: запирающая зона, или плотное соединение (zonula occludens), зона (поясок) сцепления (zonula adherens) и пятно сцепления (macula adherens).
Запирающая зона (плотное соединение) – участок, где наружные поверхности прилегающих плазматических мембран сливаются и образуют поясок шириной 0,1–0,5 мкм вокруг всего апикального периметра клеток.
На препаратах, приготовленных с использованием замораживания – скалывания, анфас этой зоны имеет вид разветвляющейся и анастомозирующей сети внутримембранных нитей и бороздок на соответствующих поверхностях клеток.
Возрастающая сложность и число нитей (в апикально-базальном направлении) определяют плотность соединения.
Функция: предотвращает поступление (или выход) веществ из полости органа в межклеточные промежутки, т. е. внутрь стенки органа, тем самым обеспечивая защиту внутренней среды организма от проникновения из содержимого кишки различных болезнетворных агентов (бактерии, кокки и т. д.).
Зона сцепления (промежуточное соединение, опоясывающая десмосома) – второй компонент соединительного комплекса. Он простирается по всему периметру соединяющихся эпителиальных клеток чуть ниже (базальнее) от запирающей зоны.
Призматические мембраны клеток в этой зоне утолщены, образуют пластинки прикрепления, которые содержат актинсвязывающие белки: α-актинин, винкулин, плакоглобин – и находятся на расстоянии 10–20 нм друг от друга.
Прилежащие к плазматическим мембранам участки цитоплазмы клеток укреплены сетью из актиновых микрофиламентов, которые простираются до запирающей зоны.
Межклеточный промежуток часто содержит аморфный или волокнистый материал, в состав которого входит адгезивный трансмембранный гликопротеин Е-кадгерин, обеспечивающий связь между клетками.
Функция: связывает и структурно поддерживает запирающую зону.
Пятно сцепления (десмосома) – третий компонент соединительного комплекса, но часто находится и в других участках цитолеммы, соединяя эпителиоциты друг с другом.
Это дисковидное соединение между соседними эпителиальными клетками. Прилежащие клетки отделены промежутком шириной от 15 до 25 нм, а на внутренней поверхности каждой из соседних призматических мембран имеется дисковидное уплотнение (пластинка прикрепления) диаметром около 0,5 мкм и толщиной 12–15 нм, содержащее особые белки: десмоплакины, плакоглобин и десмокальмин.
Промежуточные (диаметром менее 10 нм) кератиновые микрофиламенты (тонофиламенты) из цитоплазмы клетки проникают в эти дисковидные уплотнения и выходят из них, образуя петли.
Межклеточное пространство между двумя половинками десмосомы содержит материал низкой электронной плотности и тонкие нити – трансмембранные адгезивные белки (десмоколлины и десмоглеины), которые, как считается, стабилизируют это соединение.
В центре межклеточной щели часто содержится линейное уплотнение (центральная линия).
Щелевидное соединение (нексус) – это дисковидный специализированный участок, состоящий из определенного числа субъединиц (коннексонов) – трансмембранных структур диаметром 9—11 нм.
Такое соединение типично не только для эпителия, но и для клеток ЦНС, сердечной мышцы и гладких мышц (в них получило название «нексус»).
Противоположные плазматические мембраны разделены щелью шириной 2–3 нм, но соединены коннексонами. Каждый коннексон включает шесть (иногда 4–5) цилиндрических субъединиц (состоят из белка коннексина), расположенных радиально вокруг центрального канала (1,5 нм в диаметре).
Коннексоны противоположных плазматических мембран простираются в межклеточную щель и выстроены таким образом, чтобы допустить проход ионов и мелких молекул (мол. масса не более 1200) из клетки в клетку.
Латеральные интердигитации – пальцевидные или неправильной формы выпячивания цитоплазмы боковых поверхностей соседних клеток, соединяющие друг с другом эпителиальные клетки. За счет интердигитаций (зубчатых или пальцевидных) увеличиваются прочность соединения эпителиоцитов друг с другом и площадь поверхности, через которую могут осуществляться межклеточные обменные процессы.
Базальная поверхность эпителиальных клеток прилежит к базальной мембране, к которой она прикреплена с помощью полудесмосом.
Базальная мембрана (пластинка) – неклеточная поддерживающая структура (толщиной 20—100 нм), продуцируемая лежащим на ней эпителием; видна под электронным микроскопом.
Функции: обеспечение прочной связи эпителия с подлежащей соединительной тканью, избирательная фильтрация питательных веществ, поступающих в эпителий, поддержка нормальной поляризации и дифференцировки эпителия, регуляция его роста и движения при репаративной регенерации.
Базальная мембрана образована главным образом коллагеном IV типа, ламинином и протеогликанами. Состоит из трех зон: менее плотной светлой пластинки (lamina lucida) толщиной 30–50 нм, прилежащей к плазматической мембране, более плотного сцепления волокон – плотной пластинки (lamina densa) толщиной 50–60 нм и ретикулярной пластинки.
В глубь светлой пластинки от полудесмосом эпителиоцитов направляются тонкие якорные филаменты.
Светлая пластинка содержит гликопротеины (в том числе ламинин), протеогликаны (гепарансульфат).
В плотную пластинку вплетаются якорные фибриллы (из коллагена VII типа), которые соединяются с коллагеновыми фибриллами подлежащей соединительной ткани. Плотная пластинка содержит коллаген IV типа, энтактин, гепарансульфат, адгезивный гликопротеин (фибронектин).
Ретикулярная (фиброретикулярная) пластинка состоит из коллагеновых фибрилл соединительной ткани, связанных с якорными фибриллами. По толщине значительно превосходит светлую и плотную пластинки. Фибриллы образованы коллагеном I и III типов.
Полудесмосома выглядит как половина десмосомы. Присутствует в области базальной мембраны клеток некоторых видов эпителия, таких, как клетки базального слоя многослойного плоского эпителия, миоэпителиальные клетки молочной и слюнных желез, клетки базального слоя эпителия трахеи. Прикрепляет клетки к подлежащей базальной мембране.
Складки базальной плазмолеммы – специализированные образования, типичные для эпителия, транспортирующего ионы. Формируют глубокие впячивания (базальный лабиринт), разделяющие митохондрии.
Полагают, что ионные насосы (Mg+, К+, АТФ) в плазмолемме находятся в тесной связи с источниками энергии (АТФ из митохондрий).
Апикальная поверхность эпителиальных клеток образует разнообразные выпячивания (микроворсинки, стереоцилии, реснички и жгутики).
Микроворсинки – пальцевидные выросты эпителиоцитов (около 1 мкм в длину) диаметром 0,1 мкм, выступающие в просвет органа и увеличивающие его всасывающую поверхность. Они формируют щеточную каемку проксимальных частей почечных канальцев и всасывающих клеток (энтероцитов) тонкой кишки.
Характеризуются наличием гликокаликса (углеводная оболочка) на наружной поверхности. Он сформирован в первую очередь разветвлениями концевых олигосахаридов мембранных интегральных белков и фосфолипидов. Содержат пучок из около 30 актиновых филаментов (продольно ориентированных), проходящих от сердцевины микроворсинки к терминальному сплетению апикальной части цитоплазмы эпителиоцита.
Стереоцилии – длинные микроворсинки, достигающие в длину 5–7 мкм, способные к ветвлению. Присутствуют в придатке семенника и семявыносящем протоке внутренних мужских половых органов, участвуя в процессах всасывания продуцируемой ими жидкости.
Реснички – способные к активному движению специализированные органеллы (см. рис. 3.4) определенных видов эпителия (например, трахеи, бронхов, маточных труб и т. д.), которые транспортируют вдоль своей поверхности различные вещества (частицы пыли, сажи, слизи и т. п.). Имеют длину от 5 до 10 мкм и диаметр порядка 0,2 мкм.
Каждая ресничка в своей основе содержит аксонему, которая состоит из 9 пар (дублетов) микротрубочек, кругообразно расположенных вокруг двух лежащих в центре микротрубочек (система 9+2). Центральная пара микротрубочек окружена центральной оболочкой, от которой к периферическим дублетам расходятся радиальные спицы.
В каждом дублете микротрубочки связаны друг с другом мостиками из белка нексина. От каждого из дублетов трубочек в стороны отходят два ряда «ручек».
«Ручки» состоят из обладающего АТФ-азной активностью белка динеина, который расщепляет АТФ для энергетического обеспечения движения ресничек.
В основании каждой реснички находится цилиндрическое базальное тельце, по строению идентичное центриоли (также построено по принципу 9+0 из 9 триплетов микротрубочек, расположенных радиально, как спицы в колесе).
Единственный тип клеток человека, имеющих жгутик – спермии. Они содержат только по одному жгутику длиной 50–70 мкм и имеют сходное с ресничкой строение.