



Станислав Дробышевский
Достающее звено. Книга 2. Люди
Очень древние Homo: Homo rudolfensis
После 2 млн лет остатки Homo становятся более частыми. Правда, ясности от этого не прибавляется, число вопросов, как обычно в таких случаях, только растет. Бóльшая часть находок сделана в Восточной Африке.
Особенно много окаменелостей обнаружено в Кооби-Фора – одной из главных палеонтологических сокровищниц Кении. Преархантропы датируются здесь временем примерно от 2 до 1,6 млн лет назад. Вместе с тем здесь же мы наблюдаем и самое большое разнообразие, с трудом поддающееся интерпретации. Практически все находки в разное время и разными авторами диагностировались и как Homo rudolfensis, и как Homo habilis, и как Homo ergaster, и как специфические виды. Иногда разница взглядов объясняется исследованием разных частей находок. Классический пример – KNM-ER 1590a-q, находка, включающая фрагменты свода черепа, верхнюю челюсть и набор зубов детеныша предположительно мужского пола 5,2–5,3 лет. Ее датировка – одна из самых больших в Кооби-Фора, порядка 1,85 млн лет назад. Округлость и грацильность свода, а также отсутствие мегадонтии позволили определить KNM-ER 1590 как Homo habilis, тогда как абсолютные и относительные размеры бугорков моляров – как Homo rudolfensis (например: Grine et al., 2009). Предполагаемый объем черепа – 870 см³ – тоже великоват для хабилисов.
Ряд других находок в Кооби-Фора часто идентифицируется как Homo rudolfensis. Некоторые из них близки по возрасту к KNM-ER 1470: 1,91–1,95 млн лет назад для верхней челюсти KNM-ER 62000 и фрагмента нижней челюсти KNM-ER 62003. Другие имеют датировки около 1,89 млн лет назад, но с уточнением возраста туфа KBS, под которым они залегают, их возраст может быть пересмотрен в сторону удревнения. Это, например, нижние челюсти KNM-ER 1482, KNM-ER 1801, KNM-ER 1802. Все они характеризуются схожим комплексом черт: большей грацильностью в сравнении с парантропами, но массивностью – с Homo habilis и H. erectus, суженностью альвеолярной дуги спереди, но широким расхождением назад, широкой и очень высокой восходящей ветвью, относительно большими молярами, не достигающими, впрочем, значений массивных австралопитеков, смесью признаков микроанатомии зубов Paranthropus и Homo.
Гораздо полнее черепная крышка с частью лица KNM-ER 3732 (все те же 1,88–1,90 млн лет назад). Промежуточные между Homo rudolfensis и Homo habilis размеры позволяют предположить, что это либо самка первого вида, либо самец второго. Другие же исследователи считают, что KNM-ER 3732 является древнейшим представителем вида Homo ergaster, поскольку он достаточно похож на KNM-ER 3883 (Schwartz et Tattersall, 2000b). Все же отличия от эргастеров показательны: KNM-ER 3732 меньше и грацильнее их, с менее развитым надбровным торусом, с объемом мозга около 700 см³, тогда как у эргастеров – около 800 см³.
Некоторые характерные черты Homo ergaster, возможно, формировались в недрах популяций эогоминин довольно рано. Хороший тому пример – затылочная кость KNM-ER 2598, чья датировка, возможно, приближается к 1,9 млн лет назад: значительные толщина, преломленность и развитие рельефа позволили диагностировать ее как принадлежащую Homo ergaster. Впрочем, учитывая данные из Дманиси, мы можем убедиться, что первые эогоминины вовсе не обязаны отличаться какой-то особенной грацильностью свода, так что нет особых оснований не включать KNM-ER 2598 в Homo rudolfensis. В челюсти KNM-ER 3734 с близкой датировкой разные исследователи также усматривают черты Homo rudolfensis или Homo ergaster.
Гораздо фрагментарнее череп KNM-ER 3891, несколько более молодой – около 1,8 млн лет назад. Он тоже был определен как Homo rudolfensis, в пользу чего свидетельствуют сильное развитие височной линии и сильный альвеолярный прогнатизм. Однако скуловой отросток верхней челюсти KNM-ER 3891 не слишком высок, альвеолярная дуга широкая и округлая спереди, а зубы не мегадонтны. Таким образом, можно видеть усиление черт, приписываемых обычно Homo habilis в более позднее время.
Рудольфенсис возвращается! Рудольфенсис возвращается?
Проблема изменчивости и разнообразия “ранних Homo”, видимо, еще долгие годы будет терзать умы палеоантропологов. Две противоборствующие концепции никак не могут одолеть друг друга. Согласно первой, среди первых представителей рода Homo имелась довольно большая половая или же межпопуляционная вариабельность. Согласно второй, наблюдаемые различия слишком велики и отражают существование как минимум двух видов: более мелкого Homo habilis и более крупного Homo rudolfensis. Установлению истины посвящены многие статьи и главы в умных книгах, мудрыми учеными применялись заковыристые математические анализы, оценки полового диморфизма и сравнения с изменчивостью обезьян и гоминид от австралопитеков до современных людей. Ничего не помогает. Как обычно, в конце таких работ звучит вечное упование на грядущие находки, которые таки прольют свет на темную загадку антропогенеза.
И вот они, эти новые находки!
В 2007, 2008 и 2009 годах в Кооби-Фора в Эфиопии были обнаружены три фрагмента черепов. Добрые антропологи подобрали им круглые номера – все любят красивые циферки с нуликами, которые и запоминаются легко. Наиболее полный и важный обломок – KNM-ER 62000 – почти полная верхняя челюсть. KNM-ER 60000 – практически целая нижняя челюсть. Наконец, KNM-ER 62003 – небольшой, но информативный кусок нижней челюсти. Датировки фрагментов – 1,91–1,95, моложе 1,87 и 1,90–1,95 млн лет назад соответственно. Сравнение этих находок с наличными коллекциями показало, что наилучшим образом они вписываются в рамки проблематичного вида Homo rudolfensis.
Еще в 1972 г. Б. Нгенео нашел череп KNM-ER 1470. Сочетание крупных почти ортогнатных челюстей, заметно приплюснутых спереди, довольно большого мозга (большого сравнительно с австралопитеками и известными тогда Homo habilis, а не сапиенсами, как иногда в запале невежества пишут креационисты) и приличной датировки (которая сама по себе долгое время была предметом ожесточенных дебатов, а ныне устаканилась на 2,03 млн лет назад) сделало проблематичным отнесение KNM-ER 1470 к какому-либо из известных тогда видов гоминид. Подвешенное состояние порешил В. П. Алексеев в 1978 г., когда в своей великой книге описал вид Pithecanthropus rudolfensis (Алексеев, 1978). Правда, он сделал это не в полном соответствии с канонами и правилами международной номенклатуры, за что до сих пор подвергается нападкам критиков. Кроме того, книга была написана на русском языке, неведомом мировой общественности. В 1986 г., однако ж, вышел английский вариант книги В. П. Алексеева, а потому несклонные вникать в исторические детали зарубежные антропологи всегда пишут Homo rudolfensis Alexeev, 1986, чем раздражают и веселят наших антропологов. Но мы-то знаем правду: Homo rudolfensis Alexeev, 1978!
Проблема, впрочем, не в бюрократических мелочах. Главной сложностью оказалось то, что долгое время Homo rudolfensis оставался видом, известным по одному-единственному черепу. Конечно, были попытки приписать туда же и другие находки, но все они были крайне фрагментарны, а потому образ не очень клеился.
Челюсть KNM-ER 62000 замечательным образом напоминает лицо черепа KNM-ER 1470. Она столь же ортогнатна и уплощена, передняя сторона ее альвеолярной дуги так же ровна, отчего дуга становится, собственно, не дугой, а буквой “П”. Замечательно, что носовое отверстие KNM-ER 62000 узкое и почти треугольное, что является типичной чертой “ранних Homo” в целом и черепа KNM-ER 1470 в частности. Зубы KNM-ER 62000 вопреки крупным размерам самой челюсти не слишком велики в сравнении с прочими “ранними Homo”; кроме того, они относительно вытянуты продольно (мезиодистально на одонтологическом диалекте).
Нижняя челюсть KNM-ER 60000 уникальна среди челюстей “ранних Homo” своей сохранностью. Удивительно, что полностью сохранились восходящие ветви – огромные, совершенно вертикальные, с эффектно развернутыми углами. Впечатляет и высота симфиза, на котором при некоторой фантазии можно даже углядеть зачаток подбородочного выступа. Как и на верхней челюсти KNM-ER 62000, альвеолярная дуга – не дуга, а прямоугольник. Собственно, именно эта черта позволила исследователям объединить KNM-ER 60000 и KNM-ER 62000. Зубы же их различаются, поскольку на KNM-ER 60000 они относительно укорочены, а не удлинены.
Примечательно, что форма альвеолярной дуги отличает KNM-ER 60000 от канонических Homo habilis. Другое дело, что бы это значило? Действительно ли отличия пропорций зубов и альвеолярных дуг свидетельствуют о видовом статусе отличий? Имеющиеся цифровые данные, строго говоря, указывают скорее на уклонение более крупных форм от мелкого большинства.
Можно обратить внимание и на более широкий набор находок. Вообще-то, форма нижней челюсти KNM-ER 60000 весьма напоминает таковую челюсти из Дманиси D2600, которая была не так уж давно описана в качестве самостоятельного вида Homo georgicus. Огромная величина восходящих ветвей и симфиза D2600 назывались в качестве эксклюзивных признаков эндемичного вида. Однако ж другие челюсти из Дманиси представляют спектр широкой изменчивости, сравнимый с размахом вариаций челюстей из Кооби-Фора. Весьма различается и форма лица дманисцев – опять же не в большей степени, чем восточноафриканские хабилисы и рудольфенсисы. Равным образом среди людей современных рас можно найти гораздо больший спектр вариаций. Правда, для этого придется сравнивать людей из разных экологических зон, зато на стороне ископаемых находок – интервалы в сотни тысяч лет.
Помнится, долгий и нудный подсчет размеров и форм челюстей и прочих обломков из Восточной Африки, проведенный в масштабе изменчивости всех гоминид, привел меня некогда к выводу, что хабилисы и рудольфенсисы расходятся лишь на расстояние подвидов (текст диссертации можно найти на сайте www.antropogenez.ru). Именно так выглядят и графики, отражающие размеры зубов новых находок в Кооби-Фора. Любопытнее, что часть восточноафриканских находок – KNM-ER 1801, KNM-ER 1802, KNM-ER 1805, UR 501 – я тогда посчитал принадлежащими новому виду массивных австралопитеков, конвергентному с “ранними Homo”. Одним из ключевых признаков этого потенциального вида была повышенная массивность в сочетании с прямоугольной формой альвеолярной дуги. Знакомое сочетание! Не его ли мы видим на KNM-ER 60000? Замечательно, что размеры зубов этих индивидов – KNM-ER 1801, KNM-ER 1802, UR 501 – занимают промежуточное положение между Paranthropus boisei и “ранними Homo”, а KNM-ER 1805 почти совпадает с KNM-ER 60000 и KNM-ER 62000. Чехарда таксономических определений продолжается…
Датировка 1,9 млн лет назад позволяет предположить, что к виду Homo rudolfensis относятся тазовая и бедренная кости KNM-ER 5881 из Кооби-Фора. Они описаны как специфические и отличающиеся от восточноафриканских Homo erectus. Собственно, отчего бы им быть похожими на эректусов, если эректусы имеют гораздо меньший возраст? По измерительным же характеристикам кости KNM-ER 5881 закономерно занимают промежуточное положение между австралопитеками и более поздними “ранними Homo”, а из последних ближе всего к хабилису OH 62.
К виду Homo rudolfensis с некоторой вероятностью относятся обломки длинных костей левой ноги KNM-ER 1481a-d, найденные также под туфом KBS и имеющие датировку около 1,89 млн лет назад. Эти кости определяли и как “раннего Homo sp.”, и как Homo habilis, и как Homo ergaster, и как Homo erectus. Бедренная очень похожа на бедренную современного человека, маленькая и грацильная. Вероятно, у многих исследователей срабатывает стереотип: раз рудольфенсисы имеют бóльшие размеры челюстей и зубов в сравнении с хабилисами, то и их посткраниальный скелет должен быть крупнее. На самом деле такого закона в природе нет; например, синхронные рудольфенсисам дманисцы имели столь же крупные зубы, но очень низкий рост и маленькие кости. Замечательно, что коленный сустав KNM-ER 1481b выглядит прогрессивнее, чем у афарских австралопитеков AL 333-x26 и AL 333–4, но практически не отличается от варианта современного человека.
Ту же датировку имеет крайне фрагментарный скелет KNM-ER 3735. Его обычно определяют как Homo habilis, но основания этому крайне зыбки. От скелета осталось довольно много обломков, но каждый из них очень невелик. Малые размеры тела, как и в случае с KNM-ER 1481, вряд ли могут сами по себе служить диагностическим признаком. KNM-ER 3735, вероятно, обладал шимпанзеподобным соотношением длин лучевой и плечевой костей – даже более примитивным, чем у Люси, – и, возможно, увеличенной длиной рук относительно ног (Haeusler et McHenry, 2007). Пропорции KNM-ER 3735 могли быть примитивнее, чем у более позднего, но менее крупного олдувайского хабилиса OH 62 (Haeusler et McHenry, 2004). Вместе с тем на височной кости KNM-ER 3735 имеется шиловидный отросток – прогрессивный гоминидный элемент, который, впрочем, изредка встречается и у австралопитеков.
Еще хуже сохранность скелета KNM-ER 1500a-p, имеющего все ту же датировку 1,89 млн лет назад. Несмотря на наличие фрагмента нижней челюсти, видовая принадлежность остается спорной – Paranthropus boisei, Australopithecus sp., Homo rudolfensis или Homo habilis. Множество примитивных признаков сочетается на этих фрагментах с более-менее прогрессивными. Рост, как и в предыдущих случаях, был небольшим.
Обычно как Homo ergaster или даже Homo erectus определяется правая тазовая кость мужчины KNM-ER 3228. Однако ее стратиграфическое положение – ниже туфа KBS – определяет датировку более 1,87 млн лет назад, возможно 1,95 млн лет назад. Учитывая, что в этих же слоях встречены черепа и зубы только Homo rudolfensis, гораздо логичнее предположить, что и таз принадлежал особи этого вида. Замечательно, что пропорции таза и места прикрепления мышц на нем существенно отличаются от типичных для австралопитеков и приближаются к современному варианту. Кость крупная, так что рост мог достигать даже 1,8 м.
Одна из самых молодых находок, определяемая как Homo rudolfensis, – нижняя челюсть KNM-ER 60000, имеющая датировку 1,78–1,87 млн лет назад (Leakey et al., 2012). Она же является и самой целой из относимых к этому виду. Невозможно не отметить ее крайнего сходства с челюстью D2600 из Дманиси, что особенно впечатляет, учитывая их синхронность. Кстати, и размеры неба более древней находки KNM-ER 62000 в наибольшей степени совпадают с дманисскими D2282 и D2700, по ширине – и с D4500.
Впрочем, небольшой фрагмент тела нижней челюсти KNM-ER 819 тоже очень массивен, а потому иногда также определяется как Homo rudolfensis. Между тем его новейшая датировка – всего 1,5 млн лет назад (McDougall et al., 2012). Таким образом, либо вид Homo rudolfensis продержался дольше, чем обычно считается, либо есть какая-то погрешность в датировке, либо челюсть относится на самом деле к Paranthropus boisei, либо вся концепция самостоятельности Homo rudolfensis неверна, либо, что вероятнее всего, KNM-ER 819 отражает начало увеличения массивности челюстей у нового вида Homo erectus.
Несколько сотен тысяч лет существования вида Homo rudolfensis – длинный и недостаточно изученный этап нашей эволюции. Эти люди жили не там, где их кости могли успешно сохраняться, они еще не хоронили умерших и совершенно не заботились о потребностях будущих антропологов. Парантропы, обитавшие ближе к воде, известны намного лучше. Только упорные усилия современных исследователей могут пролить свет на эти “серые тысячелетия” нашей предыстории.
Не самые древние Homo: Homo habilis
В слоях выше туфа KBS в Кооби-Фора размеры костей эогоминин уменьшаются, а потому практически всегда их определяют как Homo habilis. В частности, к ним относятся фрагменты нижних челюстей KNM-ER 1501 и KNM-ER 1502, чья датировка порядка 1,6 млн лет назад. Более полные находки из поздних слоев Кооби-Фора – KNM-ER 1805 и KNM-ER 1813 – специфичны, а потому будут рассмотрены позже.
Классические Homo habilis найдены в Олдувае. Они не синхронны и достаточно разнородны, но, к сожалению, крайне фрагментарны: не обнаружено ни одного целого черепа. Находки гоминид сопровождаются примитивными галечными орудиями, отчего вся культура получила название олдувайской. К древнейшим эогомининам Олдувая относится маленький фрагмент нижней челюсти OH 4 с датировкой 1,9 млн лет назад. Сохранившийся второй или третий моляр большой, но все же меньше, чем у Homo rudolfensis из Малави и Кооби-Фора.
Строение эогоминин Олдувая времени порядка 1,79–1,86 млн лет назад известно по черепу OH 24, или Твигги, предположительно женскому. Обычно он считается образцовым представителем вида Homo habilis, но малые размеры и некоторые черты строения позволили ряду авторов отождествить его с Australopithecus africanus, другие же видят сходство с Homo rudolfensis. Все же ряд параметров позволяет отличить OH 24 от австралопитеков: это умеренное заглазничное сужение, поднятие затылочного отдела, вертикальное расположение барабанной пластинки, другие детали строения височной кости и основания черепа. Вместе с тем отсутствие шиловидного отростка – выраженно примитивный признак, лицо крайне уплощено и даже вдавлено, резко выступает вперед, носовые кости очень узкие, а по передней стороне лица идут вертикальные “передние лицевые валики”, аналогичные тем, что характеризуют южноафриканских австралопитеков и Stw 53. Все же скулы выдаются не так сильно, как у австралопитеков. Характерно строение верхней челюсти OH 24: небо очень короткое и почти квадратное, линии передних и заклыковых зубов ровные, сходящиеся под прямым углом. При этом нет никаких признаков заклыковой мегадонтии, а передние зубы относительно задних крупны. Объем мозга 560–590 см³, то есть больше, чем у австралопитеков. Строго говоря, чисто морфологически OH 24 вполне может быть расценен как австралопитек, но очень и очень продвинутый, фактически это очередное “достающее звено” между Australopithecus и Homo. Кстати, очень похожи на Australopithecus africanus и другие олдувайские гоминиды близкого возраста, например OH 68 и OH 70.
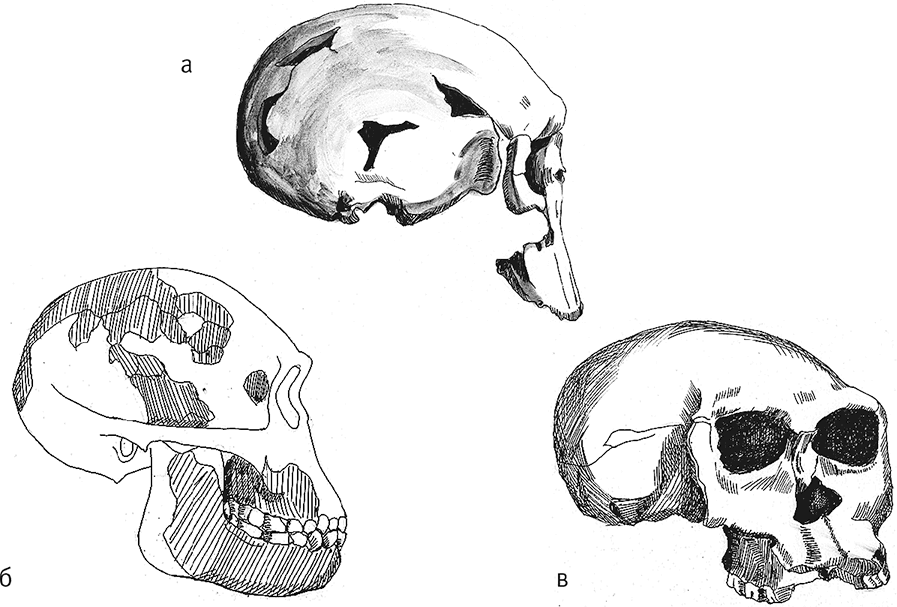
Рис. 13. Череп KNM-ER 1470 (а), черепа OH 13 (б) и OH 24 (в).
Довольно сильно от OH 24 отличается верхняя челюсть OH 65, имеющая аналогичную датировку. Ширина и высота OH 65 несколько меньше, тогда как размеры неба, альвеолярной дуги и зубов – заметно больше, чем у OH 24. Посему ряд антропологов склонны сближать OH 65 с эогомининами типа KNM-ER 1470, а OH 24 – с мелкими вроде KNM-ER 1813 (например: Clarke, 2012).
Голотип вида Homo habilis – обломки двух теменных костей, нижняя челюсть и кости кисти OH 7 – имеют датировку 1,74–1,82 млн лет назад (в 2006 году еще один зуб из той же челюсти был найден в вышележащем слое, так что ранее определявшийся возраст мог быть завышен; Leakey et al., 1964). Не исключено, что кости принадлежат разным индивидам. Забавно, что длина реконструированной теменной кости довольно велика, несмотря на предположительно подростковый возраст, почему некоторые авторы склонны сближать OH 7 с KNM-ER 1470 в противовес мелким индивидам, предположительно представляющим другой вид (Rightmire, 1993); в этом случае смысл термина Homo rudolfensis теряется, а часть хабилисов перестают быть хабилисами. Казалось бы, в пользу этой точки зрения говорит и большая датировка OH 7, однако теменная кость мелкого KNM-ER 1813 в реальности длиннее, чем у большого KNM-ER 1470, а OH 7 занимает между ними промежуточное положение, тем более что реконструкция OH 7 довольно условна. Предполагаемый объем мозга был 657–680 см³. Нижняя челюсть OH 7 не может быть названа массивной, так что ее отождествление с Homo rudolfensis Кооби-Фора и Малави спорно, но подростковый возраст не позволяет делать четких выводов.
Человек ли человек умелый?
Что и говорить, вопросов относительно первых Homo немало. Антропологи неустанно ведут жаркие споры о статусе как целых видов, так и отдельных находок. Примером такой битвы за истину может служить очередная статья, опубликованная недавно в журнале Nature (Spoor et al., 2015). В ней авторы явили на суд ученой аудитории новые реконструкции теменных костей и нижней челюсти OH 7 – голотипа вида Homo habilis. Проблема в том, что нижняя челюсть хабилиса сломана и сплющена, а теменные кости разбиты и почти не смыкаются друг с другом. Вновь реконструированная нижняя челюсть получилась весьма примитивной, с вытянутой альвеолярной дугой. Авторы считают, что она оказывается примитивнее, чем даже на полмиллиона лет более древний Homo AL 666–1 из Хадара (это, правда, верхняя челюсть, но, имея одну, можно понять форму и противоположной). Авторы также аргументируют, что Homo habilis резко отличается от Homo rudolfensis, так что они не могут представлять один вид.
Теменные кости OH 7 реконструировались неоднократно, но отчего бы не сделать это еще разок, благо в лаборатории простаивают томограф да ядреный компьютер? И вот очередное цифровое колдовство свершилось, и на глаза удивленной публики выплывают цифры 729–824 см³! А ведь это в полтора раза больше, чем считалось доселе! Как же это объяснить? Авторы реконструкции считают, что все логично: просто большой мозг эволюционировал раньше и активнее, чем форма челюстей.
Если суммировать итоги реконструкции челюсти и черепа, получаются интересные выводы: Homo habilis – это не предковый для современного человека вид, а тупик эволюции, настоящим же предком был эфиопский AL 666–1. Эволюция мозговой коробки была более стандартной и ранней, тогда как челюсти менялись замысловатее; Homo habilis сохранял примитивный вариант долго после появления гораздо более продвинутых людей. Впрочем, и Homo rudolfensis – хоть и ближе к нам, чем Homo habilis, но тоже тупик.
Вот такие пироги!
Как обычно, есть пара-тройка но.
Во-первых, форма альвеолярной дуги – штука до крайности изменчивая. Хотелось бы узнать – каких современных людей авторы брали за образец? А ведь среди особо прогнатных восточных экваториалов – австралийских аборигенов и меланезийцев – частенько встречаются индивиды с прямоугольной зубной дугой, резко вытянутой, с клыками в выраженных углах между спрямленной передней линией резцов и прямыми параллельными линиями заклыковых зубов. Что, их тоже теперь относить к австралопитекам? Политкорректность не треснет?
Во-вторых, реконструкция объема мозга по раздолбанным теменным костям – дело неблагодарное. Доселе для OH 7 публиковался разброс вероятных размеров мозга от 560 до 724 см³. В. И. Кочеткова реконструировала целые эндокраны OH 7 в трех вариантах, и ведь самый крупный из них оказался как раз самым неправдоподобным! “Австралопитековый” имел размер всего 560 см³, а “умеренный” – 650 см³. Другие авторы предполагали величину около 687 см³. Но 800 см³ – не многовато ли? Томография и компьютеры – это, конечно, здорово, но классику ведь тоже забывать не стоит. Да и есть ли смысл убиваться по поводу размеров именно этого черепа – они всегда останутся спорными, – если есть несколько почти целых черепов: OH 24, KNM-ER 1470, KNM-ER 1805 и KNM-ER 1813.
В-третьих, спор о принадлежности “ранних Homo” к одному или нескольким видам ведется уже полвека, и никому пока не удалось победить. Еще одна реконструкция старой находки вряд ли способна поставить окончательную точку в этой грандиозной битве мозгов.
В-четвертых, если H. habilis – тупик, H. rudolfensis – тупик, то кто же тогда предок? (На этом месте должны появиться креационисты и радостно закричать: “АГА!!!”) Авторы статьи считают, что H. erectus, но относят к этому виду дманисцев, так что выходит, что эректусы совершенно синхронны и хабилисам, и рудольфенсисам. Точка зрения до крайности спорная.
Кажется, путаницу и противоречия можно успешно разрешить, если больше внимания уделять хронологии и не смешивать группы, отстоящие друг от друга на сотни тысяч лет. Тогда будет видно, что H. rudolfensis – более древний и массивный вид, включающий в себя в том числе дманисцев, H. habilis – более молодой и грацильный, а H. ergaster и тем более H. erectus – это уже совсем другая история. Такая схема не избавляет от необходимости выискивать тупики эволюции, но в ней наблюдаемая изменчивость приобретает эволюционный смысл, а количество непонятных ветвей резко сокращается.
Так что интрига сохраняется, битва за первого человека в самом разгаре, впереди еще много интересного! Как говаривала Алиса: чем дальше, тем любопытственнее и любопытственнее…
Особый интерес представляет кисть OH 7. По ряду признаков она может быть определена как специализированная – в частности, своеобразную форму и очень маленькие размеры имеет ладьевидная кость. Существенно, что сустав между костью-трапецией и первой пястной костью был седловидный – “трудовой”, хотя и сильнее уплощенный, чем у современного человека. Примитивное строение имеет IV запястно-пястный сустав. Основание II пястной кости по некоторым параметрам ближе к горилльему варианту, чем человеческому, а сама кость довольно грацильна. Соотношение широтных размеров пястных костей OH 7 больше напоминает обезьяний вариант и очень редко встречается у современного человека. На фалангах продольные борозды головок и борозды для сгибателей пальца на телах очень глубокие, а валики по бокам фаланг бугристые, так что тела фаланг расширены в средней части; все эти признаки скорее как у человекообразных обезьян, нежели человека. Вместе с тем головки концевых фаланг расширены, что является одним из важнейших признаков трудовой кисти. Особенно велика концевая фаланга I пальца кисти, хотя она может относиться к стопе, другому индивиду или даже другому виду. Совокупность признаков кисти свидетельствует, что 1,7–1,8 млн лет назад кисть еще не обрела всех человеческих свойств; вместе с тем это самая человеческая кисть из древнейших, намного более человеческая, чем была у австралопитеков.
Такая же мозаика примитивных и прогрессивных черт обнаружена на стопе OH 8, найденной тут же; не исключено, что ей обладал тот же индивид. С одной стороны, стопа принадлежала, очевидно, полностью прямоходящему существу и имела хорошо выраженные продольный и поперечный своды, с другой – своеобразна, а по ряду черт равно отличается от понгид и современного человека (Kidd et al., 1996). В частности, очень оригинальны таранная, пяточная, ладьевидная и кубовидная кости, большой палец, вероятно, был сильнее отведен, чем обычно у современного человека, но при этом первая плюсневая крайне массивна, да и другие длинные и мощные и, вероятно, чуть более подвижные, чем у человека. Таким образом, стопа хабилисов еще могла сохранять адаптацию к жизни на деревьях, но в силу ли эволюционной инерции или действительно использования по этому назначению – неясно. Справедливости ради надо сказать, что ярлычок с большого пальца стопы, если он и был, не сохранился, так что видовая принадлежность OH 8 вообще-то точно неизвестна. Некоторые антропологи считают, что это стопа бойсовского парантропа. Вопрос может быть окончательно решен только обнаружением более-менее целого скелета парантропа или хабилиса.
В слоях с датировками 1,7–1,8 млн лет назад или чуть больше вместе с останками Paranthropus boisei были найдены мелкие фрагменты черепа с большой и малой берцовыми костями OH 6, а также большая и малая берцовая кости OH 35. Судя по всем указанным посткраниальным костям, рост олдувайских эогоминин был невелик, порядка 1,6 м.
Не только размеры, но и пропорции можно установить по остаткам скелета взрослого индивида OH 62, известным также как Дик Дик Хилл или Ребенок Люси и имеющим датировку порядка 1,8 млн лет назад. Скелет состоит из 302 фрагментов, но в данном случае, к сожалению, “много” не значит “хорошо”. Все же можно понять, что по большей части признаков OH 62 соответствует другим мелким Homo habilis Олдувая и Кооби-Фора. Впрочем, и тут не обошлось без альтернативных мнений: отдельные исследователи склонны считать, что OH 62 больше похож на Australopithecus afarensis, чем на Homo habilis (Berillon et Marchal, 2002). В отличие от OH 24, этот гоминид не имел “передних лицевых валиков”, так что нет оснований предполагать его родство с южноафриканскими австралопитеками. Гораздо интереснее все же его посткраниальные особенности. Размеры костей небольшие, так что рост достигал всего 1–1,25 м. Существенно, что руки относительно ног были очень длинными, даже длиннее, чем у Australopithecus afarensis (Richmond et al., 2002). OH 62 и KNM-ER 3735 представляют два самых изученных скелета “ранних Homo”, и в обоих случаях пропорции конечностей выглядят весьма архаичными; судя по ним, эволюция от четвероногих приматов к людям не была совсем прямой, имела место специализация пропорций в первой половине “загадочного миллиона”.
В скором будущем мы узнаем о строении хабилисов намного больше, так как в Кооби-Фора найден скелет KNM-ER 64062 с датировкой 1,82–1,86 млн лет назад (Jungers et al., 2015). От него сохранились кости рук и ног. В предварительной публикации анонсирована смесь примитивных и прогрессивных черт в их строении, кажется с преобладанием именно прогрессивных.
Из несколько более поздних слоев Олдувая – 1,7 млн лет назад – происходит фрагментарный череп подростка OH 16, или Джорджа. Таксономически он определялся как Homo rudolfensis, Homo habilis, Homo ergaster, Homo sp. nov. или ранний Homo erectus, а первоначально даже как Zinjanthropus boisei. В принципе, OH 16 может считаться образцовым хабилисом; он мало отличается от OH 24. Размеры этого черепа оказываются самыми усредненными из известных для гоминид “загадочного миллиона”, в частности, объем мозга составляет 638 см³, типичны для хабилисов и детали строения височной кости. Надбровные дуги выступают несколько сильнее, чем у большинства хабилисов и рудольфенсисов, но слабее, чем у эргастеров и тем более эректусов. Примитивными чертами являются близкое схождение височных линий и очень большие размеры зубов, такие же, как у австралопитеков; особенно велики клыки.
Примерно то же можно сказать о находке OH 13 (Синди, или Синдерелла): как и OH 16, она определялась всеми возможными способами – Homo habilis, Homo sp. nov., Homo ergaster, Homo erectus. Ее возраст колеблется от 1,65 до 1,78 млн лет назад (Spoor et al., 2007). В действительности фрагменты свода черепа с наибольшей вероятностью принадлежат взрослой особи, а верхняя и нижняя челюсти – подростку. Размеры черепа, челюстей и зубов вполне соответствуют типичным для Homo habilis. Впрочем, сообразно поздней датировке, OH 13 обладает и прогрессивными чертами, например скругленностью альвеолярной дуги спереди и расхождением ее ветвей назад. Вместе с черепом и челюстями были найдены фрагменты плечевой и лучевой костей, свидетельствующие о малых размерах тела OH 13.




