



Станислав Дробышевский
Палеонтология антрополога. Том 3. Кайнозой
Инге, Володе и Маше – моей любимой семье
* * *
Мы живём в холодном мире. Причина этого проста – наша Солнечная система дряхлеет. Солнце щурит свои постаревшие протуберанцы, а Земля ёжится, теряя былой задор и покрываясь сединой полярных шапок. Всё глубже железо, всё медленнее вращается в недрах ядро.
Давно минули времена младенческих капризов и непостоянства – метеоритных бомбардировок и бурных извержений докембрия. Забыты невинные детские забавы – строительство кубиков-строматолитов, неумелые каракули на их срезах, и лишь в чулане ради ностальгии пылятся последние замки, на которые иной раз можно взглянуть, с улыбкой поминая, как это было когда-то весело. Брошены бесчисленные беспозвоночные игрушки кембрия, ордовика и силура, яркие фантики девона и карбона, подростковые – «как у взрослых» – увлечения перми – темноспондилы и зверообразные. Миновали кризисы взросления – метания триаса в поисках самоопределения, вымирания в его начале и конце: «Эх, никто меня не понимает, начну всё сначала!» Прошла и зрелость – сто сорок уравновешенных миллионов лет юры и мела, с более чем серьёзными неспешными делами, величественными масштабами, однообразным и повторяющимся, но необходимым бытом, со сложными, но лаконичными и проверенными конструкциями динозавров и птерозавров, наряду с изобретением изощрённых изысков вроде цветковых растений, эусоциальных насекомых и плацентарных млекопитающих.
Настала старость. Альбомы с чёрно-белыми ретушированными фотографиями напоминают о безвозвратно ушедшем, на полочках хранятся осколки прошлого и сувениры из прожитого. На смену лихим авантюрам, смелым экспериментам и безрассудным приключениям приходит мудрость – планета начинает осознавать себя и Вселенную, появляется Человек…
Введение
Книга, в чтение которой Вы, Уважаемый Читатель, уже погрузились, – третий том «Палеонтологии антрополога». Первый был посвящён докембрию и палеозою, второй – мезозою. Нелишним будет в самом начале повторить некоторые существенные моменты.
Автор книги – антрополог, специалист по ископаемым людям. А палеонтология – наука о самых разных живых существах прошлого и, более того, условиях их существования. Значит ли это, что антрополог не может рассуждать о бронтотериях и гиенах, крокодилах и фороракосах? Многим так и кажется. Однако настоящий антрополог как раз обязан попытаться разобраться во всём разнообразии былых экосистем, ведь именно в хитросплетениях отношений водорослей и рачков, магнолий и сумчатых, копытных и грызунов, а всех их вместе взятых – с нашими предками – и формировались наши человеческие качества. Экологические взаимосвязи бывают весьма неожиданными, без их изучения появление новых признаков у предков кажется произвольным и порой даже бессмысленным. Понятно, что многие части биосферы на наше происхождение не особо-то и влияли, но для того, чтобы понять – влияли или нет – с ними тоже надо бы разобраться. Вот и приходится антропологу вникать в сложности жизни планктонных сообществ и влияния климата на облесённость материков.
Ясно, что постичь все тонкости всех частных биологических дисциплин невозможно – слишком обширна современная наука, слишком много данных она накопила. Очевидно, что трудно объять необъятное. Но попробовать-то можно! А Читатель, следя за приключениями предков, возможно, уловит и какие-то новые закономерности, иные взаимосвязи – тех хватит ещё не на одно поколение исследователей.
Понятно, что в формате галопа по миллионам лет неизбежны упрощения и сокращения, не исключены искренние заблуждения и даже ошибки в сложных и спорных вопросах. Невозможно помянуть все точки зрения и все нюансы прошлого. С чем-то некоторые специалисты будут спорить (это проверено на двух предыдущих томах, комментируя которые, разные знатоки выдавали порой диаметрально противоположные претензии) – и более чем обоснованно. Но изложенные в книге данные подкрепляются работой сотен палеонтологов на протяжении пары сотен лет. Часть этих трудов процитирована в конце книги; все перечислить невозможно, иначе число страниц превысило бы все разумные пределы. К тому же, в науке постоянно происходят какие-то удивительные открытия, так что эта книга может служить ориентиром в бесконечном обилии информации, а Читатель при прочтении сможет направить более пристальный взор в заинтересовавшую его сторону.
Во избежание лишних споров в книге по возможности не уточняются ранги больших таксономических групп. Для упрощения текста существа после упоминания латинского названия иногда называются русской транскрипцией. Если в роде один вид, пишется и видовое название, если видов много, указывается только родовое. Живых существ миллионы, ясно, что упомянуты далеко не все, а лишь самые примечательные, больше всех повлиявшие на нашу эволюцию или те, о которых узнал автор.
* * *
Прежде чем приступать к истории жизни, хотелось бы выразить глубочайшую признательность и высказать огромнейшее спасибо всем, кто способствовал улучшению данной книги. Мелине Ананян координировала работу над книгой с самой идеи до выхода в свет, а Юлия Лаврова – именно третий том. Александр Борисович Соколов взял на себя тяжкий труд поиска редакторов и уговорил их на подвиг быстрейшего прочтения и комментирования немаленького текста. От всей души благодарю научного редактора – Алексея Анатольевича Бондарева – за огромные усилия и потраченное время, без него многие нюансы я изложил бы ошибочно, искажённо или устарело. И конечно, заранее извиняюсь перед Алексеем Анатольевичем, что, идя на поводу популярной направленности книги, я учёл не все его желания. Многие важные с научной точки зрения моменты были опущены, многие термины, формулировки и сложные моменты заведомо упрощены во имя доступности неподготовленному читателю, так что, если Уважаемый Читатель найдёт ещё какие-то упущения, вся вина лежит исключительно на авторе.
Отдельная благодарность Руслану Зайнуллину и Евсею Рябову, в невообразимых количествах снабжающих меня новостями; многие интересные новые статьи без Руслана и Евсея я бы пропустил.
* * *
Третий том «Палеонтологии антрополога» продолжает нашу историю. Тут рассказывается о жизни кайнозоя – нашей эры, на данный момент конечной. Шестьдесят шесть миллионов лет – не так уж много для целой эры, сопоставимо с некоторыми периодами былых времён. Но сколь много значат эти миллионы для нашей эволюции! Если в предыдущие сотни миллионов лет мы раскачались лишь до состояния недоземлеройки, то в эти неполные семь десятков успели вырасти, спуститься на землю, встать на две ноги, по пути пару-тройку раз сменить диету, взять в руки палки и камни, поумнеть, заговорить и даже начать писать и читать книги про то, как мы раскачались, успели, встали, сменили, взяли, поумнели, заговорили и начали… Начали же!
Часть IV. Кайнозой: ещё не вечер?
Палеоген. 66–23,03 миллиона лет назад: от мира карликов до мира гигантов
Международная и российская шкала:
66 млн л. н. – палеоцен: датский век – 61,6 – зеландский век – 59,2 – танетский век – 56 – эоцен: ипрский век – 47,8 – лютетский век – 41,2 – бартонский век – 37,71 – приабонский век – 33,9 – олигоцен: рюпельский век – 27,82 – хаттский век – 23,03
Палеоцен. 66–56 миллионов лет назад: мир карликов
Первый период кайнозойской эры – палеоген, а его первая эпоха – палеоцен. Гигантские ящеры вымерли, а на их место заступили карлики – крошечные наследники мезозоя, сначала робко, но со всё большим энтузиазмом начавшие реставрировать пострадавшую от вымирания биосферу. Среди них совсем немаловажное место занимали наши прямые предки – первые приматоморфы.
* * *
Карта начала палеогена похожа и не похожа на современную. Без труда узнаются все континенты и архипелаги, океаны и заливы, но во всём есть какая-то странность. Европа – остров, отделённый от Азии широким морем-проливом, в окружении россыпи мелких островков. Южная Азия – вытянутый по широте архипелаг, на дне проливов которого кроются будущие вершины Гималаев. Индия и вовсе дрейфует посреди будущего Индийского океана. Аравия ещё не отделилась от Африки и никак не соединена с Азией, а поперёк Сахары досыхает огромный залив Средиземного моря. Между зарождающейся Индонезией и Австралией, только что отпочковавшейся от Антарктиды, лежит огромное море – слияние Индийского и Тихого океанов. Атлантический океан почти замкнут на севере и совсем узенький на юге, зато на месте Панамы вольготно и плавно перетекает в Тихий, а поднимающиеся Анды отделены от Амазонии длинным узким заливом Карибского моря.
Весь предыдущий меловой период на дне морей откладывался CaCO3, то есть мел, собственно, и давший последнему периоду мезозоя его название. Но время шло, тектоника неумолимо ворочала литосферные плиты, и местами – в зонах океанических разломов – рано или поздно донные отложения погружались в глубины планеты (для этого есть умный термин «субдукция»). Безграничные толщи мела плавились в магме, отчего CO2 высвобождался и поднимался в атмосферу, вызывая парниковый эффект и приводя к потеплению в общепланетарных масштабах. Так в отдалённой перспективе сработало наследие водорослей кокколитофорид, когда-то снизивших парниковый эффект и тем заморозивших динозавров. Ныне же, когда злые ящеры сгинули, температуры вновь скакнули вверх. Правда, на протяжении палеоцена эта тенденция срабатывала ещё не вполне однозначно, и пару раз даже намечалось похолодание, но в любом случае климат был гораздо теплее большей части и мелового, и юрского, а уж по сравнению с нынешним и подавно – аж на 10 °C курортнее! В целом жизнь однозначно налаживалась!
Местами это восстановление можно проследить очень детально. Пепелища пожаров, вызванных падением метеорита, густо поросли грибами – пресловутый иридиевый слой, отложившийся при оседании пыли от падения астероида, просто перенасыщен грибными спорами (между прочим, подобная же история намного раньше случилась на границе перми и триаса, где грибы тоже первыми встречали первые лучи мезозоя на руинах палеозоя). Кстати, часть спор могут быть и не грибными, а водорослевыми, что тоже логично: симбиоз грибов с водорослями – лишайники – универсальные выживальщики, первыми заселяющие любую пустошь, так как водорослевый фикобионт способен к фотосинтезу, то есть берёт энергию из света, а из воды и углекислого газа синтезирует органику, тогда как грибной микобионт разлагает органику обратно. Никакие консументы – существа, перерабатывающие органику в другую органику, то есть мы, – в таком раскладе вовсе не нужны, они лишни и избыточны. Через некоторое время эти грибные плантации начали снова зарастать лесами.
В колорадском Коррал Блюффс палеонтологи крайне точно датировали подробнейшую последовательность напластований с массой ископаемой флоры и фауны. Оказывается, что непосредственно после мел-палеогеновой катастрофы резко увеличилось количество папоротников, пальм, за следующие сто тысяч лет разнообразие млекопитающих удвоилось, а за триста тысяч лет размеры их тел утроились. Через семьсот тысяч лет после границы колорадщину топтали уже очень приличного размера звери, например полусоткилограммовый Eoconodon coryphaeus, отъедавшийся на свежепоявившихся бобовых. Крокодилы и черепахи дополняли картину тропического болота. Но эти снулые рептилии уже не могли остановить торжества пушистых и теплокровных зверей!
Как вы там, потомки?
Современные бобовые растения Fabacea – чудесные создания! Во-первых, у них шикарные двусторонне-симметричные цветы с широкими лепестком-парусом сверху, узкими вёслами по бокам и парой соединённых в одну лодочку – снизу. Когда пчёлка или другой какой опылитель ползёт к желанному нектару по такой конструкции, на тельце гарантированно налипает пыльца. Для этого нижние девять тычинок обычно срастаются в желобок и намазывают пыльцу на пузико пчёлке, а одна свободная тычинка аккуратно, но неумолимо шлёпает сверху, прилепляя пыльцу на голову и спину. Опыление при таком раскладе просто неизбежно!
Вторая, куда более важная особенность бобовых – клубеньки на корнях, домики для клубеньковых азотфиксирующих бактерий Rhizobiales. Эти бактерии выбирают из атмосферы несвязанный азот и заключают его в состав органических молекул. А это процесс не такой простой. Азота-то в атмосфере у нас навалом, а вот связывать его умеют немногие, при том, что азот входит в состав кучи необходимых всем органических соединений, включая нуклеиновые кислоты. Наличие бобовых на любом луге несказанно увеличивает плодородие почвы, так как все прочие растения тоже могут получить связанный азот на дармовщинку.
Появление бобовых неизбежно должно было мощно двинуть эволюцию растительных экосистем. Оно и двинуло, планета начала зарастать густыми лесами – нашим домом на долгие последующие миллионы лет.
* * *
Морские экосистемы кайнозоя восстанавливались едва ли не быстрее наземных. Сильнее всего поменялись самые нижние и самые верхние экологические уровни – планктон и хищники. Полностью исчезли все хищные морские рептилии – плезиозавры и мозазавры. Шанс на захват океана получили хрящевые рыбы – химеры, акулы и скаты. Любопытно, что в конце мела сгинули лишь две их большие группы – Hybodontida и Synechodontiformes, причём вторые, во-первых, всё же обнаруживаются в палеоцене, а во-вторых, могли даже не вымереть, а стать предками для некоторых современных хрящевых. Однако недостатки их дыхания – отсутствие жаберных крышек – и системы поддержания равновесия – отсутствие плавательного пузыря – видимо, уже были неисправимы. Конечно, и сейчас повстречаться с акулой в открытом море – сомнительная радость (вернее, радость, но только для акулы), а несколько позже – в эоцене и далее – они даже дали новое разнообразие, но свой шанс хрящевые так до конца и не реализовали.
Без особых проблем пережили катаклизм осетры, а вот костные ганоиды сохранились лишь в виде реликтов. Из позднего мела в палеоген и вплоть до современности переходят роды осётров Acipenser и панцирных щук Lepisosteus, с палеоцена известны и амии Amia.
На этом безрыбье костистые рыбы вдруг оказались полновластными царицами морей. Мы ведь сейчас живём на планете рыб – число их видов и жизненных форм намного больше, чем всех прочих позвоночных, вместе взятых.
Конечно, и им позднемеловое вымирание аукнулось более чем основательно. Огромное число линий пресеклось, а после из остатков быстро заново сформировались новые формы. Частично они повторяли то, что уже плавало в мезозое.
Из всего этого великолепия особое внимание обращает на себя Serrasalmimus secans из палеоцена Марокко. Его родичи были весьма колоритны и более разнообразны до вымирания, но и выживший потомок не подвёл: его челюсти были плотно уставлены страшными треугольными зубами, практически как у современных пираний. Скорее всего, сходство это не обусловлено родством, а конвергентное, то есть объясняется приспособлением к одинаковым условиям.
* * *
Без особых проблем преодолели вымирание амфибии, черепахи и крокодилы. Те же роды, тот же темп исчезновения и появления новых видов: астрономия и геология, все эти ваши астероиды и вулканы, этих существ явно не волновали.
Хуже пришлось ящерицам и змеям: в фаунах Северной Америки исчезло пять шестых видов; правда, тут же несколько и появилось. Показательно, что пропали самые крупные твари – весом более полукилограмма. Значимое разнообразие чешуйчатые восстановили уже в благословенном эоцене. Впрочем, размеры росли быстрее изменчивости: в конце палеоцена колумбийский гигантский удав Titanoboa cerrejonensis достиг 13 м и веса более тонны. Это круче даже многих мезозойских ящеров. Позвонки современных удавов – отнюдь не маленьких, трёх с половиной метровых – выглядят на фоне позвонков титанобоа просто крошечными. Такие монстры явно должны были и питаться кем-то соответствующим. Сопутствующая фауна включает двоякодышащих и костистых рыб, черепах и специфических крокодилов Dyrosauridae. Последние – Anthracosuchus balrogus, Acherontisuchus guajiraensis и Cerrejonisuchus improcerus, – видимо, были главной добычей титанической змеи. Между прочим, здешняя черепаха Carbonemys cofrinii – тоже рекордсменка: её панцирь был 1,72 м в длину! Что там было подмешано в воду, что их так пёрло?! Весь этот серпентарий бултыхался в жарких тропических болотах Южной Америки под покровом протосельвы – прототипа нынешних джунглей. Млекопитающих там, кстати, не замечено, хотя в принципе на материке они в это время были. Получается, век гигантских рептилий в Южной Америке с концом мезозоя вовсе не закончился! И лишь спустя некоторое время звери и тут взяли своё.
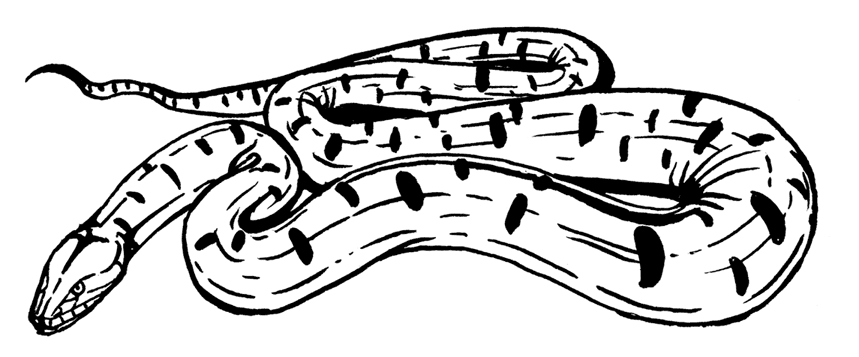
Titanoboa cerrejonensis
Как часто бывает, исчезновение одних привело к появлению других: среди чешуйчатых рептилий возникли амфисбены Amphisbaenia. Древнейшими представителями были Oligodontosaurus wyomingensis, Archaerhineura mephitis и Chthonophis subterraneus из Вайоминга, а также Polyodontobaena belgica и Camptognathosaurus parisiensis из Бельгии и Франции. Их челюсти, плотно усаженные толстыми зубами, очень уж характерны.
Как вы там, потомки?
Современные амфисбены Amphisbaenia чаще рассматриваются как подотряд чешуйчатых, но иногда предлагается выделить их в самостоятельный отряд, настолько они своеобразны. В отличие от ящериц и змей, тело амфисбен покрыто сплошной роговой оболочкой, подразделённой поперёк и немножко вдоль, отчего внешне амфисбены почти неотличимы от дождевых червей. Кожа слабо соединена с телом, так что легко сдвигается, позволяя амфисбенам двигаться едва ли не перистальтически, причём как вперёд, так и назад, что, во-первых, объясняет альтернативное название «двуходки», а во-вторых, ещё больше делает их похожими на червей.
Но достаточно посмотреть на череп, чтобы стало ясно: амфисбены – вовсе не мирные червячки! Череп амфисбен максимально похож на череп «чужого» из известного фильма: вытянутый, массивный, почти безглазый, со страшными зубастыми челюстями (что довольно странно, учитывая питание термитами, муравьями и дождевыми червями; впрочем, вспоминая кротов и землероек, может, не так уж и странно), а вместе с формой тела – на червей из «Дюны»[1]. Головой амфисбены копают, так что форма заточена под это дело, причём у разных родов и видов реализованы вариации роющих инструментов: под плоскую лопату, под заострённую мотыгу, под кол или отбойный молоток, причём рабочий передний край может быть опущен или курносо поднят. Амфисбены могут ползать и по-змеиному, и изгибаться по вертикали, а также поднимать гибкий хвост, не отличающийся от головы, для отвлечения внимания, что, кстати, даёт их очередное название – «двухголовые змеи».
Почвенный образ жизни диктует и другие особенности. Глазки у амфисбен редуцированы и скрыты под кожей, откуда едва просвечивают. Сохранилось только левое лёгкое, кстати, в отличие от змей и безногих ящериц с их одним правым, что заодно показывает независимость эволюции амфисбен. Ручки и ножки пропали, и только североамериканские Bipes сохранили крошечные передние ручки – трёх- или пятипалые.
Загадочно географическое распространение амфисбен: возникли они в Северной Америке, откуда мигрировали в Европу и Африку, а уже из Африки – сильно позже и причём дважды независимо – обратно через Атлантику – один раз прямо на Карибы и второй – туда же, но транзитом через Южную Америку. Выходит, амфисбены несколько раз успешно преодолели морские просторы, будучи почвенными роющими созданиями! После этого остаётся лишь удивляться, как же малоподвижны были наши предки!
* * *
На пернатых катаклизм сказался странно-избирательно: часть линий типа энанциорнисов и конфуциусорнисов исчезла без следа, а веерохвостые птички сдюжили. Как обычно с хрупкими пташками, при изучении древних фаун надо делать основательную скидку на плохую сохранность эфемерных косточек.
Как минимум на границе палеоцена и эоцена уже существовал отряд древненёбных птиц-палеогнат Lithornithiformes. Эти птички, например много видов Lithornis, в наибольшей степени (и внешне, и по сути) напоминали современных южноамериканских тинаму или (не по родству, а чисто внешне) – куропаток. Строение их нёба выдаёт принадлежность к древненёбным птицам, большой киль явно свидетельствует о хороших летательных способностях, а длинные ноги намекают на околоводность. Судя по мозгу Lithornis plebius, эти птицы имели отличное обоняние, хотя и уступающее современным киви, но сопоставимое с тинаму. Благодаря шикарной сохранности черепа Lithornis promiscuus, а также некоторых других родственных птиц, удалось установить, что кончик клюва литорнисов был очень чувствительным, как у современных тинаму, киви и ибисов. Такая вибротактильность, очевидно, нужна птицам, постоянно ковыряющимся в земле, так что легко представить берега североамериканских и европейских болот раннего кайнозоя, по которым среди травы снуют «курочки» и тычут длинными острыми клювиками в тину.
Как вы там, потомки?
Современные тинаму Tinamiformes – внешне невзрачные, но очень важные для эволюции птички. Они весьма похожи на куропаток, однако родственны южноамериканским страусам нанду, что особенно здорово видно по поведению. У тинаму неплохое по птичьим меркам обоняние; питаются они всем подряд. Тинаму имеют большой киль на грудине, но их грудная мускулатура белая, то есть имеет мало митохондрий и не может обеспечить долгий надёжный полёт. Поэтому тинаму предпочитают прятаться и застывать на месте, лишь в крайнем случае убегая и уж совсем в безнадёжном – взлетая. Но их коротенький хвостик не всегда позволяет лететь куда полагается, так что тинаму могут даже врезаться в коварные деревья, бросающиеся наперерез. Гнёзда у тинаму очень простенькие, а многие виды откладывают яйца просто на землю. Как и у страусов, в одно гнездо откладывают яйца несколько самок, а высиживает самец; он же потом печётся о птенчиках, которые, впрочем, весьма самостоятельны с момента вылупления. Тинаму очень уязвимы, почти все птенцы гибнут, зато и взрослеют меньше чем за месяц. Все эти особенности прямо отсылают к дремучим мезозойским временам. Неспроста тинаму сохранились лишь в Южной Америке, где бодрых плацентарных хищников не так много. Впрочем, там их не так уж мало – с полсотни видов.
Древненёбные птицы с самого начала были склонны к наземной жизни и быстро теряли способность к полёту, а уж с исчезновением хищных динозавров эта тенденция получила отличные возможности для реализации. Тут же появился целый веер нелетающих страусоподобных птиц: например, Remiornis heberti из Франции и Diogenornis fragilis из Бразилии – птицы неясного родства, но, вероятно, близкие страусам-нанду.
Древненёбные птицы, впрочем, не задались. На первое место выпорхнули новонёбные.
Ясно, что наилучшим образом известны околоводные аналоги или даже прямые родственники гусей и уток. Несколько видов Presbyornis на своих колченогих лапах вышагивали по камышам Северной Америки и Центральной Азии, вытягивали длинные шеи и собирали широким вогнутым клювом – прямиком из «Утиных историй» – всяческую тину.
Гусеобразные птицы жили и в Южном полушарии: почти полная внешняя копия пресбиорниса Conflicto antarcticus летала над самой Антарктидой, которая тогда имела вполне умеренный климат и была покрыта лесами, а Australornis isoni из Новой Зеландии был родственен меловым антарктическим Vegavis iaai и Polarornis gregorii.
В той же Новой Зеландии обнаружен и самый примитивный и одновременно самый маленький пелагорнис Protodontopteryx ruthae. Пелагорнисы Pelagornithidae – странная группа птиц, появившаяся в начале кайнозоя и вымершая совсем недавно – когда наши предки уже начинали пользоваться орудиями труда. Две черты пелагорнисов обращают на себя особое внимание. Во-первых, это были похожие на альбатросов планёры от полутора до шести метров в размахе крыльев – одни из самых огромных птиц за всю историю планеты. Во-вторых, их челюсти были какой-то зверской пилой, так как их края покрывали далеко отстоящие, но очень большие зазубрины. Это не были настоящие зубы, а именно выросты челюсти; поэтому альтернативное название пелагорнисов – «псевдозубатые птицы». Вероятно, такими клювами удобно было выхватывать из воды кальмаров и крупных, но не слишком бронированных рыб. Скорее всего, первоначальный успех пелагорнисов был связан с вымиранием гигантских морских птерозавров, а также зубатых птиц ихтиорнисов. Экологическая ниша планеров-рыбоядов пуста не бывает! Видимо, и итоговое вымирание было связано с изменениями в рыболовецкой области, скорее всего – с появлением крутых конкурентов типа современных альбатросов и чаек. Впрочем, довольно долго пелагорнисы сосуществовали с современными птицами-рыбоядами, так что конкурентная гипотеза явно не самодостаточна. Учитывая время исчезновения – 2,5 миллиона лет, то есть время мощных перестроек климата, – причина, скорее всего, хитро-экологическая, но детали процесса пока ускользают. Впрочем, это было сильно позже палеоцена…
Понятно, что рыбу можно ловить по-разному. Исчезли же не только летуны – птерозавры и ихтиорнисы, но и нелетающие гесперорнисы. Экологическую нишу прибрежных ныряльщиков тут же, конечно, заняли пингвины Sphenisciformes. Богатейший палеоценовый пингвинятник открыт в Новой Зеландии: древнейший и примитивнейший Muriwaimanu tuatahi, его чуть более поздняя и продвинутая версия Waimanu manneringi, ещё более прогрессивные Kumimanu biceae и Crossvallia waiparensis, а также иные глупые птицы робко прятали тела жирные в утёсах. Уже муривайману и вайману были размером с императорского пингвина, кроссваллия достигала роста 160 см и веса 70–80 кг, а кумиману – 160–180 см и 90–100 кг! Даже самые ранние пингвины были вполне узнаваемы: мешковидное вертикальное тело на коротких ножках, загребущие крылья-ласты, длинный острый клюв для ловли рыбы. Довольно быстро они распространились по Южному полушарию, где и процветают поныне.
Имелись птицы и попроще. Над мелководными лиманами моря Волгоградской области порхали Volgavis marina, похожие на серебристых чаек, но, возможно, более родственные фрегатам или буревестникам.
Работа над ошибками
Первоначально волгавис был датирован поздним маастрихтом, но после возраст оказался раннепалеоценовым. Кстати, в одной статье с описанием волгависа упоминается плечевая кость птерозавра-аждархида из недалёкого от волгависа местонахождения, найденный в позднепалеоценовом слое и признанный перемытым из маастрихтского. Но, строго говоря, переотложенность исходит скорее из самого факта птерозавровости. А так-то, почему бы отдельным птерозаврам и не пожить чуток в палеоцене?.. Фантазия, конечно, но, возможно, стоило бы обращать на подобные находки особое внимание?
Длинноногая птица Scaniornis lundgreni, вероятно, похожая на фламинго (что, впрочем, весьма спорно, так как остатки крайне фрагментарны), шагала по берегам палеоценовых Швеции и Германии. Целая орнитофауна границы мела и палеоцена обнаружена в формации Хорнестаун в Нью-Джерси: гусь Anatalavis rex, буревестник Tytthostonyx glauconiticus и фаэтон Novacaesareala hungerfordi. Как и в предыдущих случаях, тут найдена околоводная фауна. Что происходило в лесах и горах – большей частью покрыто мраком.
Впрочем, и про это немножко известно благодаря находкам во Франции, в местонахождении Серне-ле-Реймс и некоторых других. Самыми эффектными палеоценовыми лесными птицами были, без сомнения, Gastornis parisiensis и G. russeli – ближайшие родственники кур. Эти двухметровые нелетающие «петушки» имели толстенные трёхпалые ноги и огромадный клюв, похожий на клещи-болторезы для срезания стальных цепей и штырей. Конечно, такой колоритный облик был интерпретирован как хищническая адаптация: якобы гасторнисы были злобными притеснителями несчастных первобытных зверюшек. Образ демонической убер-курицы, терзающей панически разбегающихся протолошадок, прочно вошёл в палеогеновый бестиарий популярных книг о палеонтологии. Однако новейшие исследования показывают, что реальность, видимо, была совсем не такой. Изотопные анализы и изучение следов прикрепления мышц на челюстях показали, что гасторнисы были не плото-, а плодоядными. Получается, самый близкий аналог гасторнисов – дронт! Можно провести аналогию и с туканами или птицами-носорогами, но с поправкой на способности к полёту. Кроме прочего, это значит, что крупнейшими растительноядными существами Европы в палеоцене были птицы, то есть экосистема была во многом похожа на мадагаскарскую до прибытия туда людей. В общем-то, и географически ситуации схожи, ведь Европа была скорее архипелагом, чем материком. Кстати, до Северной Америки в палеоцене гигантские птицы ещё не добрались, так что там звери имели лучшие шансы на развитие своих вегетарианских наклонностей, которые они, собственно, успешно и реализовали. Учитывая ещё и архаичных страусов, тема больших нелетающих птиц была в палеоценовой Европе очень модной.
Если одни пернатые ограничивали наши возможности конкуренцией, другие просто нагло нас жрали! Конец палеоцена ознаменовался появлением сов – Berruornis orbisantiqui во Франции и Ogygoptynx wetmorei в Колорадо. Вот только наши предки избавились от гнёта злых динозавров, не прошло и десятка миллионов лет – как прилетела новая напасть! Думается, именно такие невзгоды были главными движителями нашей ранней эволюции: ночные хищники выдавили нас из ночного образа жизни, придали бодрости-прыгучести и сообразительности. Как знать, кабы не совушки – круглые головушки, может, так бы и до сих пор вместо нас-умничек по ветвям ползали приторможенные плезиадапиформы…
* * *
Конечно, главное достижение палеоцена – расцвет млекопитающих. Случился он не сразу, но очень быстро. Первый десяток миллионов лет кайнозоя звери были ещё очень мелкими, довольно однообразными и трудноразличимыми, но прогресс шёл небывалыми темпами. Вымирание коснулось млекопитающих как-то очень избирательно: кто-то исчез, кто-то сократился в числе и разнообразии, а кое-кто и преуспел.
Как ни странно, даже в палеоцене по углам планеты можно было найти остатних реликтов мезозоя и едва ли не палеозоя. Так, в Канаде до конца эпохи дожил Chronoperates paradoxus – симметродонт, примитивный настолько, что поначалу его приняли даже за распоследнего зверозубого-цинодонта! В принципе, современные однопроходные-яйцекладущие тоже ведь являются подобным осколком былого, даже более примитивным, чем симметродонты.
Кстати, палеоценовые однопроходные нам тоже известны, и – неожиданно! – из Аргентины, где жил родственный утконосу Monotrematum sudamericanum. Трудно судить о животном по трём молярам, но в этом случае важнее география: зоогеографически вырисовывается соединение Австралии, Антарктиды и Южной Америки в мелу и, возможно, палеогене.




