



Станислав Дробышевский
Палеонтология антрополога. Книга 2. Мезозой
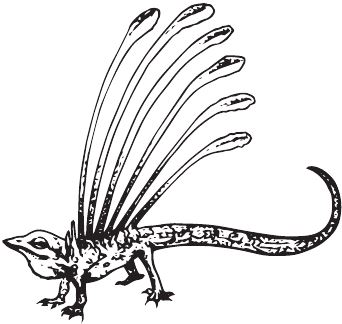
Longisquama insignis
Кроме «перьев», лонгисквама удивительна и остальными пропорциями: её треугольная глазастая головка по длине почти равна пятисантиметровому тельцу (впрочем, задняя половина скелета не сохранилась), а передние лапки – ещё длиннее, причём размеры плеча, предплечья и кисти почти одинаковы, лопатка длинная и тонкая, почти как у птиц, ключицы, вероятно, срастались в единую вилочку. Шейных и брюшных рёбер нет – вроде бы для пущей воздушности, однако кости не имели воздухоносных полостей. Всё тело было покрыто длинными армированными чешуйками.
Лонгисквама, шаровиптерикс, озимек и некоторые другие нестандартные триасовые архозавроморфы иногда объединяются в группу Fenestrasauria, хотя её статус очень шаток.
Одни из самых обычных животных триаса – Rhynchosauria. На удивление, они крайне непопулярны среди художников, существуют считанные реконструкции их облика, а на масштабных панно триасовых пейзажей Вы никогда не увидите стад ринхозавров, хотя в реальности они были аналогом современных некрупных копытных, типа свиней или овец, основным мясом для хищников, наряду с дицинодонтам. Ринхозавры были низкими коротконогими зверушками обычно меньше полутора метров длиной (впрочем, некоторые вырастали и до пяти метров), но с крупной и очень странной головой. Примитивные раннетриасовые Noteosuchus colletti и среднетриасовые Mesosuchus browni ещё имели маленький клюв с зубами, но более продвинутые Rhynchosaurus articeps и Hyperodapedon обзавелись уже здоровенным узким беззубым клювом в передней части морды. Череп непрерывно расширялся в задней части, пока у гиперодапедона ширина не превзошла длину. Задние и нёбные зубы слились в длинные и широкие зубные пластины-щётки с массой пупырышек, которыми ринхозавры перетирали жёсткие растения. Учитывая низкорослость и форму клюва с зубами, предполагается, что ринхозавры ели корни, выдёргивая их из земли. Как ни странно, активное жевание сочеталось с отсутствием вторичного нёба. Вместе с тем, наружное носовое отверстие из-за сужения передней части морды стало непарным, прямо как у млекопитающих, что крайне нетипично для рептилий.
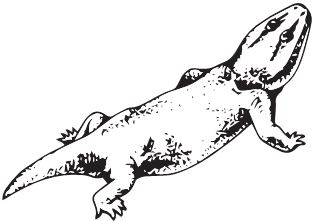
Rhynchosaurus articeps
Примечательна география ринхозавров: гиперодапедон известен в Южной Америке, Африке, на Мадагаскаре, в Индии, Северной Америке и Европе, то есть, проще говоря, везде. Неспроста в разное время разные исследователи описали массу родов и видов – Cephalonia, Isalorhynchus, Macrocephalosaurus, Paradapedon, Scaphonyx, Stenometopon и Supradapedon, которые при ближайшем рассмотрении оказываются одним и тем же гиперодапедоном.
Маленькая тонкость
Остатки ринхозавров найдены во многих местах, в том числе в Шервудской формации, названной в честь того самого Шервудского леса в Ноттингемшире, где разбойничал Робин Гуд. Знаменитый грабитель богачей не знал, что бегает по триасовым отложениям с ценнейшей палеонтологией.
В конце триаса из каких-то проторозавров появились Trilophosauria – в целом примитивные архозавроморфы, но с уникальной зубной системой. Это были приземистые тяжеловесные длиннохвостые создания 2,5 м длиной, ползавшие на раскоряченных длиннопалых когтистых лапах. Необычным было то, что у трилофозавров закрылось нижнее височное окно. У примитивного Variodens inopinatus передние зубы были коническими, а задние – резко расширенными гребнистыми трёхбугорчатыми «молярами». Дальше пошёл Trilophosaurus: его небольшая укороченная голова с высоким затылком – для увеличения жевательной мускулатуры – была снабжена спереди роговым клювом, а задние зубы стали ещё более узкими трёхвершинными гребнями, причём верхние и нижние плотно смыкались. Видимо, такая зубная система была предназначена для пережёвывания растений, причём жевательные движения происходили скорее спереди-назад, а не поперечно. Совсем странным был Teraterpeton hrynewichorum, чья морда вытянулась в длинный узкий беззубый клюв (в задней части челюстей и нёба мелкие зубы присутствовали), а ноздри растянулись на половину длины черепа.

Что сказать, триас – воистину время чудес. Но чудеса по определению не могут быть повторяющимися, слишком частыми и долгими. Да к тому же в любом сказочном лесу должны быть свои чудовища.
Особенно успешными архозавроморфами оказались собственно архозавры Archosauria. Именно они в итоге вытеснили всех и вся, в буквальном смысле съели всех удивительных родственников, да и вообще современников. В триасе архозавры пёрли напролом по принципу «всех убью – один останусь». И ведь им это почти удалось! Весь оставшийся мезозой прошёл под тяжкой пятой архозавров.
Залогом бешеного успеха архозавров стали всего две особенности: мощные зубастые челюсти с текодонтными зубами и вертикальное расположение задних ног под телом. Первое позволило кусать всех без всяких лишних заморочек с гетеродонтностью и жеванием, а второе – догнать кого угодно, чтобы его, собственно, покусать.
Маленькая тонкость
Зубы рептилий крепятся к челюстям тремя основными способами. Акродонтные зубы нарастают поверхностно или крепятся к наружной стороне челюсти, а потому непрочно; такой вариант типичен для рыб, амфибий и гаттерий. Плевродонтные зубы вырастают с внутренней поверхности челюсти, прилежа к ней на некотором протяжении, но не погружаясь в кость; они обычны у ящериц. Текодонтные погружены в специальные ячейки – альвеолы, а их основание – корень – закреплено соединительной тканью. Архозавры обрели текодонтные зубы, что дало им возможность кусать смелее и сильнее – зубы перестали вылетать по любому поводу.
Конечности у подавляющего большинства рептилий широко расставлены в стороны, так что тело большую часть времени лежит на пузике. Даже при беге пресмыкающимся часто приходится волочить брюшко и хвост по почве, что, во‐первых, дало им название, а во‐вторых, оставляет характерные изгибающиеся следы. Архозавры были едва ли не вторыми после зверообразных, кто надёжно поднялся над землёй. Только, в отличие от тероморфов, архозавры сделали ставку именно на задние ноги, а потому быстро и неоднократно стали подниматься на две ноги. Предпосылки к этому видны даже у проторозавров, для некоторых из которых, кстати, предполагается периодическая двуногость. Но архозавры решили проблему хорошо и надёжно. Бег архозавров ускорился, и никто уже не мог от них скрыться!
Современные архозавры – крокодилы – имеют едва ли не самый примитивный для архозавров вариант расположения ног. Возможно, именно архаичность позволила им пережить всех своих продвинутых родственников. Но в минуты особой бодрости (когда, например, приспичило догнать кого-то особо шустрого) даже крокодилы способны проявить свою архозавровую сущность – поднимаются на выпрямленных ногах и скачут галопом, да к тому же способны подниматься на две ноги. Правда, они делают это редко и неохотно, но наследие триасовых предков всё ещё живо в их ногах.
Особая прыткость в тёплом триасе не требовала особой теплокровности. Смотря на крокодилов, можно предположить, что уже в триасе развилось четырёхкамерное сердце и диафрагма, но на их же примере видно, что эти усовершенствования – не залог и не обязательное следствие теплокровности. Впрочем, обмен всё же не мог не ускориться. А побочным эффектом усиленного обмена и необходимостью при более шатком и сложном способе передвижения стало развитие мозга и усложнённое поведение.
Архозавры не слишком разнообразны, но это не облегчает их классификации. В классической схеме выделяются текодонты Thecodontia, крокодилы Crocodylia, птерозавры Pterosauria, ящеротазовые динозавры Saurischia и птицетазовые динозавры Orhithischia, но существует масса альтернативных схем, в которых текодонты расформировываются на кучу слабосвязанных группок, и даже динозавры то подразделяются на несколько отрядов, то перегруппировываются в совершенно невообразимом порядке. Апофеозом новаторства является включение в архозавров птиц Aves. Эти несомненные потомки архозавров по кладистической логике обязаны быть в них включены, хотя, с точки зрения орнитологов, такое объединение имеет очень мало смысла.
Маленькая тонкость
Существуют несколько способов представления родства живых существ. Самое модное и практически полностью победившее нынче направление – кладистика. Ключевая идея кладистики – классификация должна максимально отражать филогению, то есть порядок возникновения одних групп от других. Есть несколько вариантов кладистики, в самом жёстком и каноничном масштаб возникающих у потомков отличий от предков вообще не играет никакой роли, во внимание принимается лишь последовательность ветвления эволюционных линий. Соответственно, и названия можно давать лишь всем потомкам какого-то предка (монофилетической группе), но нельзя особо выделять лишь одну часть потомков, игнорируя другую. Исходя из этой логики, многие традиционные группы теряют свой смысл. Например, если птицы ответвились от некой части архозавров, а млекопитающие – от части зверообразных рептилий, то, при признании птиц и млекопитающих самостоятельными группами, нет никаких «рептилий» (поэтому многие современные зоологи очень болезненно реагируют на словосочетание «зверообразные рептилии») и даже нет динозавров (отчего другие столь же рьяно не забывают повторять «птицы – это тоже динозавры!»). При последовательном применении этой методики на планете нет даже прокариот (если признать самостоятельность эукариот от прокариот, то бактерий и архей нельзя объединять в одну группу) или эукариот (если признать прокариотами бактерий с археями, то эукариоты – потомки прокариот – получаются частью прокариот, что как бы странно); а учитывая, что эукариоты с большой вероятностью – смесь архей с бактериями, то каша получается совсем знатная. Не уверен, что читатель на этом месте не запутался, но систематика всегда была лабиринтом. Как часто бывает, при столкновении отличной идеи с реальностью приходится юлить и изобретать костыли.
Причины популярности кладизма очевидны. Он исходно имел благие и чётко сформулированные цели, строго определённый понятийный аппарат и терминологию. Он обеспечен удобными в использовании компьютерными программами, позволяющими быстро и без особых усилий строить филогенетические схемы.
Однако, как и любая идея, кладистический подход даёт сбои при слишком бездумном и фанатичном применении. Один из главных минусов – чрезмерное изобилие этих самых схем. При недостатке и разнородности исходных данных результат часто получается слишком неустойчивым. Выходит, что классификация, исходно призванная внести порядок в наши представления, становится непрерывно меняющимся калейдоскопом терминов, вводимых зачастую одноразово, «на одну статью». Всё же до установления реальной филогении, то есть происхождения и эволюционного ветвления, многих групп ещё далеко, и уже сейчас привязывать наименования групп к только лишь предполагаемым филогенетическим связям не слишком правильно.
В итоге частенько получаются не эффективные, а лишь эффектные, необычные, а потому эпатирующие схемы. К сожалению, это даёт нехороший эффект при потогонной грантовой системе, когда исследователь вынужден в минимальные сроки выдавать много-много статей. При этом одним из важнейших показателей статьи, обеспечивающим публикацию в модном высокорейтинговом журнале (и стало быть, увеличивающим шанс получить ещё один грант) является нестандартность, «вау-эффект», часто перевешивающий даже смысл работы. Если громко заявить: «Птиц нет, вокруг нас порхают динозавры!» – это произведёт намного большее впечатление, нежели долгий занудный анализ тонких особенностей строения. И не беда, что исследователь может никогда не вернуться к этой теме, получив через годик грант на совсем иную. Не беда, что квалификации при такой системе не приобретёшь. Компьютер всё посчитает, только вводи цифры.
Правда, исходные данные – те самые цифры – вводит всё же учёный. А ввести он может разный набор (даже не учитывая спорность многих показателей, связанную, например, с плохой сохранностью). Да к тому же разные алгоритмы кластерного анализа, столь любимого разработчиками программ для расчёта филогенетических деревьев, дают сильно неодинаковые итоги даже при одном исходном наборе. Всё это неизбежно приводит к бесконечной чехарде и неустойчивости схем. Но и это не беда – путаница очень хороша для обоснования необходимости новых исследований. Только вот фейерверк бывает очень утомителен…
Великий предок архозавров – Teyujagua paradoxa из самого начала триаса Бразилии. Она настолько примитивна, что у неё ещё не было предглазничного окна, хотя уже появилось отверстие в нижней челюсти. У тейюягуи были и другие странности: разомкнутое в задне-нижнем углу нижнее височное окно, непарное носовое отверстие, отверстие для теменного глаза. Зато её длинные зубы были зазубрены по краю, а что ещё надо для успешного распиливания добычи? С этого момента история мезозоя была предопределена.
Работа над ошибками
Фирменная фишка архозавров – предглазничное окно, снимающее нагрузки, приходящиеся на конец морды, и позволяющее успешнее хватать и удерживать добычу. Поскольку окно следует сразу за ноздрёй, неопытные художники могут принять его за глазницу, тогда морда резко укорачивается, а задняя часть головы резко вырастает, придавая животному гораздо более интеллектуальный вид.
В боковой части нижней челюсти у большинства архозавров тоже есть отверстие с теми же целями. Однако на не слишком грамотных рисунках и модельках черепов его регулярно забывают.
Текодонты Thecodontia, как и многие примитивные группы, быстро достигшие эволюционного успеха, классифицируются с трудом. С одной стороны, план строения у всех примерно одинаковый, с другой – в конкретных группах возникали мелкие специфические особенности, которые игнорировать тоже не получается.
В упрощённом варианте текодонты делятся на протерозухий Proterosuchia, фитозавров Phytosauria, этозавров Aetosauria и псевдозухий Pseudosuchia.
Proterosuchia – самые примитивные текодонты. Они ещё ходили враскорячку и не успели обзавестись отверстием на нижней челюсти. Их головы были большими и тяжёлыми, а передняя часть верхней челюсти загибалась крючком для лучшего заякоривания добычи. Со временем голова становилась всё тяжелее, а зубы – крупнее, хотя и реже. Примитивнейший аргентинский Koilamasuchus gonzalezdiazi мог бы быть великим предком протерозухий, если бы не жил слишком поздно для этого. Впрочем, более ранних кандидатов тоже хватает: очень примитивный китайский Fugusuchus hejiapensis, китайские и южноафриканские Proterosuchus, южноафриканские и оренбургские Garjainia, тасманийский Tasmaniosaurus triassicus и многие прочие. Образцовые протерозухии среднего триаса – Sarmatosuchus otschevi и Erythrosuchus africanus, выраставший до 5 м.
Многие протерозухии, вероятно, были водными и рыбоядными. Эта тенденция нашла законченное выражение в нескольких родственных группах.
Proterochampsia – Proterochampsa barrionuevoi, Pseudochampsa ischigualastensis и Doswellia kaltenbachi – выглядели как хорошо бронированные крокодилы, а иногда и рассматриваются как примитивная группа крокодилов. Некрупный среднетриасовый Chanaresuchus bonapartei из Аргентины и Бразилии отличался очень грацильным сложением – тонко-длинноногий, с сильно вытянутой узкой мордой, почти не защищённый, с единственным рядом пластинок вдоль спины. При всей грацильности шея у ханарезуха была короткая, мощная, с длинными рёбрами. На его стопах крайние пальцы начали уменьшаться: первый сильно укоротился, второй был толстым, но коротким, третий – нормальным, четвёртый – крайне истончённым, а пятый почти совсем исчез. По всей видимости, это была эволюция сухопутной бегательной стопы, ещё немного и могли бы получиться парно- или непарнокопытные ножки.
Другая водная группа – фитозавры Phytosauria. Это уже были завзятые рыболовы, в какой уже раз принявшие форму крокодила, тем более что они-то с крокодилами и вправду близкие родственники. Низкое длинное тело с гребнистым панцирем, короткие ножки, длинный гребной хвост, узкое длинное рыло, усаженное острыми зубами – всё узнаваемые черты. Явным отличием от своих нынешних «почти-близнецов» было расположение ноздрей не на конце морды, а перед глазами на специальном возвышении, причём хоаны открывались сразу вниз. Это позволило фитозаврам обойтись без вторичного нёба, но одновременно сдвинуть дыхательные пути назад, чтобы дышать, держа добычу во рту, да к тому же в полупогруженном в воду положении. Впрочем, как и прочие хищники, фитозавры не зацикливались на рыбалке: в желудке Parasuchus hislopi найдены два скелета мелких проторозавров Malerisaurus robinsonae. Форма рыла фитозавров могла несколько отличаться: у Belodon оно было тонкое и прямое, как у гавиала, у Smilosuchus узкое, но со вздутием сверху и загнутым расширением-утяжелителем на конце, а у Nicrosaurus kapffi – прямо-таки горбатое. Некоторые фитозавры, например, Rutiodon, вырастали до 8 м.
Работа над ошибками
Как ни странно, в списке фитозавров нет собственно самого рода Phytosaurus. Он, конечно, был описан, да вот беда – к нему относили такое количество разных видов, что в итоге он просто потерял смысл, став «родом-помойкой». Систематика фитозавров, вообще, дело запутанное.
Псевдозухии Pseudosuchia – обширная группа более продвинутых текодонтов. В широком понимании она включает массу крокодилообразных, равно как и не очень крокодилообразных тварей, в том числе и самих крокодилов (а иногда и фитозавров); в узком – только часть текодонтов, похожих на крокодилов, но не самих крокодилов.
Самая уклоняющаяся группа – этозавры Aetosauria. Возможно, они возникли из кого-то типа Revueltosaurus – метрового ящера, с виду вроде схожего со стандартными хищными текодонтами – низкого, головастого, зубастого. Вот только зубы у него были притуплённые и фестончатые по краю, явно не хищные, а растительноядные. Сами этозавры пошли ещё дальше – стали растительноядными текодонтами, от головы до кончика хвоста запаковались в плотный пластинчатый панцирь и превратились в аналог панцирных динозавров или броненосцев. В позднем триасе это было чрезвычайно актуально. Как водится, низкие, малоподвижные растительноядные существа должны иметь защиту, особенно когда вокруг шастает масса крупных хищников, и этозавры озаботились безопасностью по полной программе. Базовый примитивный вариант этозавров – некрупные Aetosaurus, чьи незамысловато бронированные скелеты иногда находят целыми кучами; очевидно, эти животные паслись немалыми стадами; у этого рода зубки ещё были острыми. Stagonolepis дорос до 3 м, его зубная система сильно поменялась: верхние зубы стали тупыми, листовидными, нижние исчезли, а передняя часть морды вздёрнулась чудным пятачком, которым можно было рыть землю в поисках вкусняшек. Панцирь из пластинчатых полос закрывал тело стагонолеписа со всех сторон, включая живот. Longosuchus meadei обзавёлся острыми треугольными шипами по бокам – от головы до конца хвоста, Typothorax вырастил шипы побольше на плечах, а Desmatosuchus вооружился огромными загнутыми шипами на шее и плечах, причём задняя пара напоминала бычьи рога. Рыло десматозуха было похоже не только на пятачок, но скорее на мотыгу.

Nicrosaurus kapffi

Desmatosuchus
Более продвинутые орнитозухии Ornithosuchia уже приподнялись на более или менее выпрямленных ногах. Предком была польская Osmolskina czatkowicensis, а несколько более продвинута южноафриканская Euparkeria capensis. Она имела самый никакой вид: мелкая помесь варана и крокодила, головастая и зубастая, ещё четвероногая, но уже, вероятно, способная подняться при случае и на две ноги. Любопытно, что Южная Африка в то время располагалась на Южном полярном круге, то есть тут были полярные ночи. Вероятно, из-за этого глазницы эупаркерии столь велики. Ornithosuchus longidens вырос уже до 2–4 м длиной и мог уверенно вставать на две ноги, хотя привычной для него всё ещё была четвероногость. Узкий и довольно высокий череп с длинными загнутыми назад зубами однозначно выдаёт злобного хищника, способного на мощный укус. От орнитозухий уже один шаг до настоящих крокодилов.
Но по пути возникло ещё некоторое количество странных боковых ветвей.
Rauisuchia – крупнейшие хищные наземные текодонты, чьи ноги совершенно выпрямились, вот только оригинальным способом: бедренная кость подходила к тазу снизу, а не сбоку, как это позже реализовалось у динозавров и млекопитающих. Базовый вариант рауйзухий – трёхметровый Ticinosuchus ferox, вроде бы крокодил, но с несколько укороченной мордой и на вертикальных ногах, покрытый сверху крупными чешуями.
Больше всех – до семи или даже девяти метров – вырос Saurosuchus galilei из Ишигуаласто. Это был самый мощный наземный хищник своего времени, превосходивший герреразавра – крупнейшего динозавра той же фауны – не только длиной, но и массивностью.
Работа над ошибками
Вероятно, именно тицинозуху или очень близкому животному принадлежат следы ихнорода Chirotherium – пятипалого существа, чьи ноги шли в линейку, как у млекопитающих, а не врастопырку, как у рептилий. Первоначально хиротерий и был описан как млекопитающее – не то медведь, не то сумчатое, не то вообще человекообразная обезьяна.
Некоторые близкие родственники рауйзухий стали очень похожи на динозавров, но всё же они не динозавры. Четырёхметровый Poposaurus gracilis отлично передвигался на двух ногах; по нему всю группу иногда называют Poposauroidea. Например, Effigia okeeffeae внешне почти не отличается от орнитомимусов: двуногая двухметровая длинношеяя, да к тому же беззубая, с широким полукруглым клювом и крошечными передними лапками, растительноядная животина. В целом так же выглядел и Sillosuchus longicervix, только его шея была ещё длиннее, а сам он вырастал до десяти метров.
Четвероногие попозавроиды – Ctenosauriscus koeneni и Arizonasaurus babbitti – обзавелись гребнями на спине из остистых отростков позвонков, как ранее это сделали платигистриксы и пеликозавры, чтобы лучше регулировать свою температуру. Аризонозавр, судя по зубастому черепу, был хищником. У Lotosaurus adentus гребень был не слишком высоким, зато в укороченных челюстях исчезли зубы, так что животное, скорее всего, было растительноядным. Таким образом, ситуация с пеликозаврами – хищными диметродонами и растительноядными эдафозаврами – практически полностью повторилась, но спустя миллионы лет и в другой группе рептилий.
Другой попозавроид – трёхметровый китайский Qianosuchus mixtus – вернулся в воду. Его острозубая морда вытянулась, хвост превратился в мощное весло. Впрочем, сильные вертикальные ноги позволяли кианозуху неплохо бегать и по земле.

Среди обилия водных и полуводных архозавров нашлось место и настоящим крокодилам Crocodylia (или Crocodylomorpha, иногда, при исключении самых примитивных групп, крокодилы определяются как отряд Mesoeucrocodylia, тогда они появились лишь в юре). Их примитивные представители – Sphenosuchia – были не слишком похожи на тех крокодилов, к которым мы привыкли. Gracilisuchus stipanicicorum вырастал примерно до полуметра и вполне мог бегать на двух ногах. Hesperosuchus agilis и Terrestrisuchus gracilis были стройными длинноногими существами с ногами-палочками, несущими щуплое тельце с длинной шеей и изящной вытянутой головкой. Неспроста в их названиях тема утончённости и грации звучит не переставая.
Последнее звено между крокодилами и динозаврами – Teleocrater rhadinus из среднего триаса Танзании – с несколькими родичами образует группу Aphanosauria. Эти трёхметровые стройные полностью четвероногие текодонты имели очень длинную шею и небольшую голову, что, впрочем, не мешало им на охоте.
Наконец, мы стремительно приближаемся к последним общим предкам самых знаменитых групп – птерозавров, динозавров и птиц. Иногда их объединяют в группу Ornithodira, иногда в Avemetatarsalia.
Достоверные прямые предки птерозавров и птиц до сих пор не найдены. Но кандидатов хватает. Один из лучших – позднетриасовый шотландский Scleromochlus taylori. Это было крошечное – 18 см – существо с относительно большой головкой, очень длинными палочкообразными ручками и ножками и очень длинным хвостиком. Судя по пропорциям ножек, склеромохлус мог бегать и на четырёх, и на двух ногах.
Птерозавры Pterosauria появляются вдруг и неожиданно в конце триаса. Понятно, что от столь хрупких, буквально воздушных, созданий осталось очень мало. Самые древние останки птерозавров обнаружены в Нью-Мексико в отложениях второй половины карнийского века, лишь чуть более поздние найдены в Аризоне и Техасе, но они столь фрагментарны, что определить по ним что-то невозможно. Парочка техасских челюстей определена как Eudimorphodon, но достоверность этого не слишком велика. Первые птерозавры относятся к Rhamphorhynchoidea в широком понимании этого термина (иногда придумываются другие названия для группы, но ключевой сути они не меняют).
Самый древний более или менее хорошо сохранившийся птерозавр – австрийский Austriadactylus cristatus из середины норийского века. Он уже имел все характерные черты группы: длинные узкие крылья в виде кожной перепонки, крепившейся на невероятно удлинённый безымянный палец кисти (размах крыльев 1,2 м), вытянутую острую голову с огромными глазницами, коротенькое тельце, не слишком маленькие ножки и очень длинный хвост. Череп австриядактилуса, как и у прочих птерозавров, облегчён за счёт увеличенных отверстий и пустот, фактически он составлен из перекладинок. Впрочем, первые птерозавры ещё не могли расстаться с зубами даже в целях облегчения веса; зубы, хотя бы и крошечные, были им дороги как архозавровая память. У австриядактилуса было два типа зубов: длинные узкие изогнутые саблевидные и короткие широкие листовидные зазубренные. По верхней стороне черепа австриядактилуса шёл бугристый гребешок – основание для гораздо более высокого прямоугольного гребня, который выполнял функцию руля и заодно служил при брачных демонстрациях.
Почти все самые древние хорошо сохранившиеся птерозавры – очень примитивный Preondactylus buffarinii, Carniadactylus rosenfeldi, Bergamodactylus wildi, Peteinosaurus zambelli и Eudimorphodon ranzii – обнаружены в отложениях конца норийского века Северной Италии, в Альпах, «значит тут у них гнездо». К ним можно добавить швейцарских Caviramus schesaplanensis, Raeticodactylus filisurensis и прочие близкие виды; Arcticodactylus cromptonellus порхал по просторам Гренландии, которая тогда, впрочем, была на широте нынешней Европы. Из их останков мы видим, что как минимум в это время полностью сложились и другие черты рамфоринхоидов – невысокий киль на грудине для крепления мощной грудной мускулатуры, оттопыренный удлинённый тонкий мизинец стопы без когтя, ромбовидная лопасть на кончике хвоста. Крылья были полностью развиты, разве что чуть относительно короче, чем у более поздних птерозавров; размах крыльев был от 45 см до метра. Впрочем, была у ранних птерозавров и своя причуда – непростые зубы: передние – вытянутые острые колышки, а задние – трёх- или пятибугорчатые пилообразные «моляры», причём верхние и нижние плотно смыкались. Такое строение не вполне понятно, учитывая рыбоядность птерозавров; а о последнем мы точно знаем, так как в желудке Eudimorphodon ranzii найдены кости рыбок Parapholidophorus.
В то же время – на границе норийского и рэтского веков – появляются и более совершённые летуны. Caelestiventus hanseni из Юты вырос до 1,5 м в размахе крыльев. Его череп стал высоким и состоящим совсем из одних пустот, лишь с тоненькими костными перемычками. Зубы целестивентуса делятся на три типа: на передней части челюстей они очень длинные и тонкие, на верхней челюсти – большие широкие короткие треугольные, а на нижней – крайне мелкие треугольные. Хорошая сохранность позволила изучить эндокран – слепок мозга – целестивентуса: на нём видно отличное развитие зрительных долей и особенно мозжечка, но слабое – обонятельных луковиц. Примечательно, что Юта в то время находилась заметно южнее, чем сейчас, и отличалась очень засушливым, прямо таки пустынным климатом (она и нынче не славится плодородием). Это не мешало жить тут птерозаврам, хотя, казалось бы, рыболовам в пустыне должно быть тяжко; впрочем, тяготы жизни им скрашивали цикадовые деревья, на которых можно было схорониться от конкурентов сфенозухий и первых динозавров.
Многие позднетриасовые продвинутые текодонты уже очень сильно похожи на динозавров. Например, Turfanosuchus dabanenesis из анизийского века Китая сочетает признаки эупаркерий, псевдозухий и грацилизухов, а внешне выглядит как очень примитивный теропод. Trialestes romeri из той самой аргентинской формации Ишигуаласто, где найдены и первые динозавры, конечно, сфенозухия, но очень уж динозавроподобная. Позднетриасовые Lagerpeton chanarensis и Dromomeron – предпредпоследняя стадия перед динозаврами, метровые длинноногие хищники, вроде бы двуногие. Впрочем, следы Prorotodactylus из оленекского яруса Польши и Франции в целом соответствуют параметрам кистей и стоп лагерпетонов, но явственно четвероногие.
Ещё ближе к динозаврам аргентинский Marasuchus lilloensis из карнийского века – тонко-длинно-двуногий стройный хищник, внешне уже неотличимый от хищных теропод и не включаемый в них только из-за специфических деталей строения. В частности, на стопе маразуха сохранялась пятая плюсневая кость, но пятого пальца уже не было. Близкий и живший там же и тогда же мелкий Lagosuchus talampayensis иногда выделяется в особую группу Lagosuchia, а иногда вообще не признаётся за особый род и вид из-за плохой сохранности его останков.
Последний уровень приближения к динозаврам – Asilisaurus kongwe из анизийского века Танзании, синхронный Lutungutali sitwensis из Замбии (что характерно, они же – древнейшие орнитодиры), Silesaurus opolensis из карнийского Польши и несколько других родственников. Эти двух-трёхметровые стройные недодинозаврики с вытянутой шеей, острыми зубками и клювом на конце морды были насекомоядными (это известно по остаткам жуков в копролитах силезавров). Но их потомки быстро развернулись и показали, на что способны пожиратели членистоногих, если дать им волю.

Lagosuchus talampayensis

Динозавры Dinosauria – группа, затмившая своей харизмой всех прочих ископаемых. С конца триаса началась их эпоха, их блеск и нищета.
Существует несколько кандидатов на звание самого-самого древнего динозавра. От признания того или другого из них зависит и место появления всей группы – Африка или Южная Америка.




