



Виталий Давидов
Боль в пояснице
Экзеркины и миокины
Начну с мышц, как «катализатора» обменных и репаративных процессов и самой «понятной» [на уровне шашлыков и куриных ножек] ткани организма.
Обыватели, в основной своей массе, рассматривают мышцы как орган, приводящий в движение рычаги скелета с целью выполнения определенной работы и служащие для поддержания тела/позы в гравитационном поле Земли. Также как орган, выполняющий вспомогательную насосную функцию, облегчающую микроциркуляцию крови в организме. Знают про участие в дыхании (диафрагма). Кто-то что-то слышал про участие в термогенезе. Как правило – и все… на этом познания и заканчиваются. Фундаментальная наука и клиническая медицина рассматривает мышечную систему в более широком аспекте/формате: помимо вышеперечисленных общеизвестных функций, учитывается и влияние мышечной ткани как самого крупного неспецифического эндокринного органа на обменные процессы всего организма. В общей сложности на данный момент установлено порядка 650 видов мышечных цитокинов (миокинов), вырабатываемых при мышечном сокращении/растяжении мышц. У человека описаны около 300. И если учитывать, что мышечная ткань у взрослого человека составляет порядка 40-60% от общей массы тела, то вклад миокинов в поддержание гомеостаза и здоровья человека просто колоссальный. Миокины действуют аутокринно, паракринно – например, мышца на кость, к которой прикрепляется и эндокринно – через кровь на мозг/печень/почки и т.д.
Чтобы было понятно, о чем идет речь, перечислю некоторые молекулярные соединения, которые синтезируются и выбрасываются в кровь при мышечной работе:
Цитокины и факторы роста:
Костный морфогенетический белок 1, 4
Нейротрофический фактор мозга
Гранулоцитарный колониестимулирующий фактор
Инсулиноподобный фактор роста 1, 1А, 2
Белки, связывающие инсулиноподобный фактор роста 2, 3, 4, 5, 6, 7
Интерлейкин 1β, 2, 4, 6, 7, 8, 10, 13, 17А, 25, 34
Макрофагальный колониестимулирующий фактор 1
Остеокластстимулирующий фактор 1
Тромбоцитарный фактор роста А, С
Секретируемый белок, связанный с frizzled 2, 4
Фактор стромальных клеток 1 (C-X-C мотив хемокин 12), 2
Трансформирующий фактор роста β (1,2,3)
Фактор некроза опухоли α
Фактор роста эндотелия сосудов A, C, D
и др.
Белки внеклеточного матрикса:
Гепарансульфат базальной мембраны
Белок ядра протеогликана (Perlecan)
Бигликан
Коллаген I (α1, α2), II(α1) и др.
Декорин
Фибриллин 1, 2
Фибулин 1, 2, 5, 7
Матриксный Gla протеин
Мимекан (остеоглицин)
Периостин
Протеогликан 4
Фактор фон Виллебранда типа А, эпидермальный фактор роста и пентраксин доменсодержащий белок 1 (SVEP1)
и др.
Энзимы:
Ферменты матриксной металлопротеиназы 2, 9, 14, 19
Супероксиддисмутаза
Активатор плазминогена тканевого типа
И прочее:
Семафорины 3(А, Б, В, D, Е), 4(В, С), 5А, 6(А, В), 7А
Рецептор эфрина типа А 1, 2, 4, 7
Экзостозин 1, 2
Белок 1 WNT1- индуцируемого сигнального пути
и др.
Биохимики и лаборанты с легкостью расширят список.
С точки зрения физической реабилитации полноценное восстановление тканей (соответственно и функции) требует меньше медикаментов и больше адекватной двигательной активности, поскольку репарация и ремоделирование поврежденной ткани всецело зависит от гомеостаза окружающих тканей, который в свою очередь зависит от общего миокинового и цитокинового фона; по-старому – от гуморального фактора мышечной работы (Голдштейн, 1961); по-новому – от синтеза «экзеркинов». Другими словами – физическая реабилитация влияет на ткани через прямую активацию/влияние на сигнальные пути. Баланс миокинов, остеокинов, адипокинов и других цитокинов во многом определяют общий гомеостаз организма и предрасположенность к заболеваниям. Миокины запускают или тормозят различные биохимические процессы в клетках, тем самым стимулируя и поддерживая полноценную работоспособность всего организма. Действия многих миокинов хорошо изучены и систематизированы.
Общие функции миокинов
Контроль механизма восстановления тканей
Контроль над воспалительным каскадом
Влияние на механизм клеточной адгезии
Анатомо-структурное развитие организма
Регуляция ангиогенеза
Влияние на иммунный ответ хозяина
Контроль секреции инсулина
Пролиферация и миграция сателлитных клеток
Организация внеклеточного матрикса
Ремоделирование внеклеточного (экстраклеточного) матрикса
Положительная регуляция локомоции
Модуляция дегрануляции тромбоцитов
Регуляция окисления жирных кислот
Противоопухолевый защитный механизм
Ниже дана краткая характеристика основных активных веществ, синтезируемых при мышечном сокращении, и описана их роль в поддержании гомеостаза организма. Привожу оригинальные названия синтезируемых «веществ», упоминаемых в медицинской и реабилитационной литературе.
Коактиватор 1- альфа гамма (γ) -рецептора-активатора пролиферации пероксисом (PGC-1α).
Свое название получил по первой открытой функции – первоначально PGC -1α был обнаружен в бурой жировой ткани как белок, взаимодействующий с PPAR γ и участвующий в адаптации термогенеза в ответ на холод.
Ключевыми регуляторами обмена липидов являются пероксисомы – это клеточные структуры, участвующие в окислении жирных кислот, синтезе липидов, холестерина, обезвреживающие токсичные вещества, а также выполняющие множество других функций.
Нарушение обмена липидов (избыток свободных жирных кислот в крови и накопление метаболически активной жировой ткани в брюшной полости и других органах) являются источником хронического неконтролируемого системного низкоуровневого (низкогредиентного) воспаления, которое может стать причиной развития различного рода хронических заболеваний (атеросклероз, артериальная гипертензия, сахарный диабет 2-го типа и т.п.).
Пероксисомы активируются специфическими рецепторами (Peroxisome proliferator-activated receptor (PPAR) при помощи транскрипционных коактиваторов PGC-1. Рецепторные белки (PPAR) служат так называемыми липидными датчиками организма, при активации которых изменяется метаболизм углеводов и липидов. Также белки PPAR являются транскрипционными факторами, т.е. белками, регулирующими экспрессию целого ряда генов, участвующих в обмене углеводов и липидов, в воспалительных и других процессах, протекающих в организме.
Поскольку метаболизм и обменные процессы в организме во многом зависят от физической нагрузки, то вполне очевидно, что именно самая активная метаболическая ткань будет задавать основной тон в данных процессах. Как раз синтез коактиватора PGC-1α при мышечной работе и будет во многом определять метаболический профиль клеток организма.
Экспрессия и активность коактиватора -1α (PGC-1α) жестко контролируются несколькими сигнальными путями, которые объединяют эндокринные события и биоэнергетические состояния. Сигналы окружающей среды, такие как воздействие холода, физические упражнения, качество и объем питания и другие «метаболические» стрессы, активируют эти сигнальные пути через катехоламины, глюкокортикоиды, глюкагон и цитокины. Клеточный энергетический стресс также активирует передачу сигналов путем изменения синтеза аденозинтрифосфата (АТФ), кислорода, активных форм кислорода (АФК) и уровня цитоплазматического Ca2+.
PGC-1α – обеспечивает прямую связь между внешними физиологическими стимулами и регуляцией митохондриального биогенеза. Является мастер-регулятором энергетического обмена. Различные типы (изоформы) PGC-1α координируют окислительный метаболизм и активируют широкую тканенезависимую программу экспрессии генов, которая управляет митохондриальным биогенезом и усиливает клеточное дыхание (регуляция синтеза транспортных белков, которые доставляют топливо в митохондрии, и антиоксидантных белков, которые нейтрализуют кислородные радикалы, образующиеся при усиленном оксидативном дыхании и пр.).
Регулярные физические упражнения стимулируют выработку PGC-1α в скелетных мышцах, что позволяет им адаптироваться к окислительному (оксидативному) стрессу, таким образом предотвращая преждевременную гибель клеток из-за повреждений митохондрий. Взаимодействуя с ядерным рецептором PPAR-гамма, определяет дальнейшее взаимодействие этого белка со многими факторами транскрипции, отвечающими за выживаемость клетки.
PGC-1α является основным фактором, который регулирует детерминацию типа мышечных волокон. Синтез PGC-1α индуцируется упражнениями на выносливость. PGC-1α регулирует метаболизм лактата, предотвращая повышение его уровня при упражнениях на выносливость; делает лактат более эффективным источником энергии.
Синтез PGC-1α стимулируется холодовым воздействием, что влияет на адаптивный термогенезом через секрецию β-аминоизомасляной кислоты при тренировке мышц.
PGC-1α ингибирует активность NF-κB – основного провоспалительного сигнального пути, подавляя продукцию провоспалительных цитокинов.
PGC-1α (изоформа 4) блокирует сигнальный каскад, активируемый Миостатином, предотвращая распад мышечных структурных белков, одновременно стимулируя гипертрофию волокон за счет воздействия на передачу сигналов инсулиноподобного фактора роста-1 (IGF1).
PGC-1α принимает участие в поляризациию макрофагов во 2 тип – противовоспалительный, таким образом, являясь модулятором иммунного ответа, что указывает на его роль в заживлении и репарации тканей. В скелетных мышцах PGC-1α вызывает секрецию факторов, которые модулируют активность резидентных иммунных клеток.
Секретируемые факторы, контролируемые PGC-1α, участвуют в системной коммуникации и влияют на обучение и поведение через передачу сигналов ирисина и кинуренина в мозге.
Физические упражнения в сочетании с ограничением калорийности пищи через коактиватор-1α (PGC-1α) приводят к: повышению синтеза гликогена, повышению транспорта и усилению окисления жирных кислот, повышению захвата глюкозы клетками, повышению митохондриального биогенеза, повышению ремоделирования и устранению повреждений в митохондриях, повышению липогенеза, повышению аутофагии, повышению синтеза цитокинов, отвечающих за ангиогенез, повышению способности к переключению типов мышечных волокон под воздействием физической нагрузки, повышению транскрипции и индукции генов, отвечающих за выживаемость клетки, повышению секреции миокинов, снижению синтеза провоспалительных цитокинов, управляет генами, ответственными за формирование синапсов (мест контакта нервного волокна и миофибриллы). В итоге действие PGC-1α суммарно повышает капилляризацию мышц, повышает физическую выносливость, мышечную окислительную мощность (дыхательную мощность), снижает мышечную дистрофию, предотвращает развитие атрофии, влияет на толерантность к инсулину, препятствует развитию старческой саркопении, поддерживает работоспособность нервно-мышечных синапсов, компенсирует митохондриальную дисфункцию и подавляет низкогредиентное системное воспаление.
Регулярная аэробная физическая активность приводит к повышению синтеза ядерного фактора, связанного с фактором эритроидного происхождения-2 (Nrf2). Данный фактор необходим для адаптации к физическим нагрузкам и обеспечивает защиту от повреждения скелетных мышц, вызванного оксидативным стрессом. Фактор (Nrf2) управляет программами транскрипции «антиоксидантных» генов, регулируя клеточную защиту от воздействия токсичных веществ и окислителей, таким образом препятствуя повреждению ДНК. Оказывает положительное влияние на мышцы, индуцируя биогенез и функцию митохондрий.
Миокины и остеокины, так называемые экзеркины (походное от английского глагола «to exert» – напрягаться, прилагать усилия).
Приведены функции наиболее важных (скорее хорошо изученных) миокинов и остеокинов, участвующих в перекрестном взаимодействии мышц и костей, и их влияние на общий метаболизм.
Интерлейкин – 6 (IL-6). Первый миокин, который выбрасывается в кровь при мышечной работе. Открыт в 1980 году. Цитокин с крутым нравом. Деструктивный, но с положительными свойствами. «Свой среди чужих, чужой среди своих». Свойства данного цитокина определяются источником его происхождения. Мышечный IL-6 (как миокин) крайне полезный и определяет весь метаболизм при физической нагрузке и восстановлении. Миокин мышечного гомеостаза. Если синтезируется клетками иммунной системы (моноцитами, макрофагами и др.) – IL-6 – третий в списке по крутости провоспалительный цитокин (после фактора некроза опухоли-альфа (TNF-α) и интерлейкина-1 бета (IL-1β)). После интенсивной физической нагрузки содержание мышечного IL-6 в плазме может возрастать в 100 раз (уровень растет по экспоненте), при этом постоянные тренировки/физическая активность поддерживают низкий базальный уровень данного миокина, если он вырабатывается мышцами непосредственно во время физической работы без первичной активации фактором некроза опухоли (при воспалении, например). «Острый» физнагрузочный IL-6 усиливает чувствительность к глюкозе и увеличивает ее поглощение, усиливает окисление жирных кислот, обеспечивая энергетическим субстратом работающие мышцы во время физической нагрузки. Стимулирует синтез инсулина бета-клетками, положительно влияет на их пролиферацию и защищает от апоптоза.
Как цитокин, IL-6 также может синтезироваться и жировой тканью (адипоцитами). У малоподвижных людей высокий уровень циркулирующего плазменного IL-6 положительно связан с развитием ожирения и инсулинорезистентностью. IL-6 ингибирует экспрессию субстрата рецептора инсулина 1 (IRS1) и транспортера глюкозы 4 типа (GLUT4) в адипоцитах (здравствуй диабет 2 типа!). Жировая ткань может обеспечить до 35% циркулирующего провоспалительного IL-6, что определяет его роль в хроническом низкогредиентном воспалении.
Участвует в регуляции иммунного ответа на повреждение тканей: при «острых» выбросах реализует свои противовоспалительные эффекты через подавление фактора некроза опухоли-альфа и интерлейкина-1бета – главных провоспалительных цитокинов. Через активацию синтеза IL-10 – главного противовоспалительного цитокина, влияет на поляризацию макрофагов во 2 тип – противовоспалительный/репаративный. Мышечный IL-6 подавляет действие главного воспалительного цитокина интерлейкина 1-бета (IL-1β) через стимуляцию синтеза антагониста его рецептора (IL-1Ra).
Хронический «выброс» жирового и/или иммунного IL-6, наоборот, усиливает действие фактора некроза опухоли – альфа (TNF-α) и интерлейкина 1-бета (IL-1β), являясь, таким образом, причиной хронизации заболеваний.
Как цитокин стимулирует оборот костной ткани путем регулирования резорбции кости и выработки остеокальцина и RANKL. Усиливает катаболизм мышечных белков в мышцах с высоким содержанием «жирового» IL-6 (неработающие мышцы), таким образом, являясь синергистом миостатина в плане развития саркопении. «Мышечный» нагрузочный IL-6 стимулирует мультипликацию сателлитных клеток в мышечной ткани, способствуя гипертрофии мышц.
Вместе с адреналином мышечный IL-6 мобилизируют и активируют специфические типы клеток натуральных киллеров (NK), которые подавляют рост раковых клеток.
Таким образом, «острый» мышечный IL-6 – хорошо; «хронический» жировой и/или иммунный – очень плохо.
Миостатин (Myostatin) – миокин гиподинамии – вырабатывается мышечными клетками и активно секретируется в кровь. Приводит к снижению роста мышечной ткани путем подавления пролиферации, дифференцировки миоцитов и синтеза белка. Негативно регулирует объем мышц: при низком уровне миостатина возникает выраженная гипертрофия; при высоком синтезе миостатина – атрофия и саркопения вследствие распада мышечных белков. Повышает степень рекрутирования и дифференцировки остеокластов, что приводит к резорбции костей за счет усиления остеокластогенеза и препятствия остеобластогенеза (костеобразования).
Оказывает системное воздействие на организм через нарушение углеводного обмена. Способствует адипогенезу (отложению жиров). Также может синтезироваться жировой тканью.
Синтез миостатина усиливается при лечении с использованием глюкокортикоидов, что вызывает ятрогенную потерю мышечной массы.
[Сама по себе мышечная ткань потребляет большое количество энергии, поэтому миостатин препятствует излишнему накоплению неиспользуемой по назначению мышечной массы].
Декорин (Decorin) – антагонист миостатина (прямое блокирование); секретируется миоцитами и высвобождается в кровь во время мышечного сокращения. Усиливает выработку основного антагониста миостатина – фоллистатина. Ускоряет заживление мышечной ткани и препятствует фиброзу мышц. Подавляет ангиогенез и онкогенез. Способствует формированию костного матрикса и отложению кальция.
Фоллистатин (Follistatin) – одновременно блокирует как миостатин, так и его рецептор (активин II б); анаболический регулятор роста скелетных мышц. Вместе с креатинином – любимая анаболическая добавка бодибилдеров (к слову: мышцы то растут, но сила не прибавляется).
Играет важную роль в подавлении воспаления в легких и уменьшении их фибротизации через блокирование рецептора активина –A, который индуцирует синтез провоспалительных цитокинов интерлейнина-1бета (IL-1β), интерлейкина-6 (IL-6) и фактора некроза опухолей (TNF-α).
Сукцинат (Succinate) – увеличивает синтез белков и стимулирует мышечную гипертрофию.
Ангиопоэтин 1 (Angiopoietin- 1; ANGPT-1) – сосудистый протектор.
Фактор роста фибробластов – 21 (FGF-21) – секретируется печенью, мышечной тканью, жировой тканью, поджелудочной железой. Участвует в регуляции углеводного и липидного обмена. Снижает уровень глюкозы и триглицеридов, повышает чувствительность к инсулину и усиливает липолитический ответ. FGF-21 участвует в «побурении» «белого» жира, окислении липидов и термогенезе, а также стимулирует экспрессию адипонектина в кровоток. Вырабатывается под воздействием холода и физических нагрузок. Усиливает экспрессию PGC- 1α.
Адипонектин (Adiponectin) – оказывает противовоспалительное действие на иммунную систему. В ЦНС способствует увеличению потребления пищи и уменьшению воспаления в гипоталамусе. В печени и скелетных мышцах увеличивает окисление жирных кислот и чувствительность к инсулину, улучшает функцию бета–клеток. Адренергическая бета-сигнализация; синтез индуцирован физическими упражнениями, взаимно способствует увеличению синтеза фактора роста фибробластов-21 (FGF-21), интерлейкина-15 (IL-15) и ирисина. Хондропротективный; стимулирует пролиферацию хондроцитов, повышает синтез коллагена II типа и аггрекана.
Интерлейкин-15 (IL-15) провоспалительный цитокин, который регулирует активность Т- лимфоцитов и клеток натуральных киллеров (NK). Как миокин обладает анаболической активностью и вызывает гипертрофию мышц. Стимулирует выработку сократительных белков и препятствует апоптозу миоцитов за счет предотвращения оксидативного стресса; способствует поглощению глюкозы скелетными мышцами и окислению жирных кислот. Снижает количество циркулирующих липопротеинов низкой плотности – основного переносчика холестерина в крови. Усиливает митохондриальную активность миоцитов. Отрицательно связан с ожирением: синтез интерлейкина-15 приводит к значительному уменьшению жировой массы тела за счет подавления дифференцировки предшественников адипоцитов. Повышает способность остеобластов накапливать минералы в костном матриксе; увеличивает минеральную плотность кости за счет уменьшения ее резорбции. Хондронегативный в больших количествах – повышает синтез матриксных металлопротеиназ (ММР). Отодвигает по срокам старение кожи.
Ирисин (Irisin; иногда – иризин) – PGC-1α зависимый: увеличивает объем мышц за счет активации миогенеза и способствует окислительному метаболизму в миофибриллах. Улучшает кортикальную массу костной ткани и подавляет остеокластогенез. Оказывает противовоспалительное действие в адипоцитах и макрофагах, повышает их способность к фагоцитозу, подавляет экспрессию провоспалительных цитокинов через ингибирование MAPK и NF-kB сигнальных путей. Оказывает антиоксидантный эффект на клетки. Оказывает системное влияние на различные органы. Защищает бета-клетки от апоптоза. Уменьшает пироптоз клеток (смерть «разрывом»). Стимулирует «побурение» белого жира и способствует термогенезу. Подавляет дифференцировку предшественников адипоцитов в зрелые адипоциты. Хондропозитивный; положительно влияет на дифференцировку хондроцитов.
Метеорин–подобный (Metrnl) – PGC-1α зависимый: при силовых нагрузках способствует гипертрофии мышц. Участвует в регуляции иммунного ответа. Противовоспалительный через подавление PI3K/AKT/NF-κB и антипироптозный. Кардиопротективный; усиливает ангиогенез в миокарде.
Инсулиноподобный фактор роста-1 (IGF-1). Открыт в 1957 году. Вырабатывается в печени, мышцах и костях – основной анаболический фактор в скелетно-мышечной системе. Способствует росту мышц. Стимулирует костеобразование и улучшает качество костей. Хондропротектор. Стимулятор нейротрофического фактора головного мозга (нейропротектор).
Фактор ингибирования лейкемии (LIF) принимает активное участие в регуляции развития мышц (способствует гипертрофии), регенерации и метаболизме. Регулирует работу остеокластов и остеобластов. Подавляет дифференцировку адипоцитов (жировых клеток). Защищает против кахексии.
Остеокальцин (OST; белок костного матрикса) – регулирует мышечную массу и способствует гипертрофии мышечных волокон, восстанавливает способность к физической нагрузке через перекрестную связь с интерлейкином-6 (IL-6). Регулирует метаболизм глюкозы и энергии и, косвенно, рост костей. Повышает чувствительность к инсулину.
Лиганд рецептора-активатора ядерного фактора-каппа–би (RANKL) – синтезируется костной тканью и некоторыми иммунными клетками. Регулирует накопление кальция (Ca2+) и активность фермента, отвечающего за его внутриклеточную транспортировку, влияет на функцию мышц. Активирует остеокластогенез и способствует потере костной массы.
Интерлейкин-8 (IL-8) – хемокин, который приманивает в основном нейтрофилы. Провоспалительный, но при эксцентрической нагрузке локально в мышцах увеличивает капилляризацию, что благоприятно сказывается на репарации и регенерации мышц.
Интерлейкин-10 (IL-10) – индуцирует противовоспалительные эффекты и повышает толерантность к глюкозе. Главный «переключатель» макрофагов из М1 типа (провоспалительный) в М2 тип (репаративный противовоспалительный).
Остеоглицин (Osteoglycin; OGN) – повышает захват и катаболизм глюкозы и жирных кислот, повышает циркулирующий уровень IL – 6. Повышает уровень щелочной фосфатазы, синтез коллагена I типа и остеокальцина.
Белок теплового шока-90 (HSP-90) – активирует интегрины, что позволяет клеткам связываться с ирисином.
Белок теплового шока-70 (HSP-70) – кардиопротективный.
Белок теплового шока-72 (HSP-72) – повышает чувствительность к инсулину и толерантность к глюкозе.
Белок теплового шока-27 (HSP-27) – снижает клеточный апоптоз.
Оксид азота (NO) (мышечный, нагрузочный/постнагрузочный) – вазодилататор; улучшает функцию эндотелия сосудов.
Фракталкин (CX3CL-1) – защищает бета-клетки поджелудочной железы (синтез инсулина) против фактора некроза опухоли-альфа (TNF-α). Усиливает нейрогенез в гиппокампе. Регулирует резорбцию кости, усиливает апоптоз хондроцитов.
Альфа–кетоглутаровая кислота – стимулирует мышечную гипертрофию и снижение массы жировой ткани.
Интерлейкин-7 (IL-7) – иммуномодулятор. Играет роль в развитии мышечных клеток. Регулирует резорбцию кости (подавляет остеокластогенез); хондронегативный (в больших количествах разрушает хрящевую ткань).
(Мышечный) Нейротрофический фактор головного мозга (BDNF) способствует росту мышц за счет участия в окислении жиров. Связывается с остеобластами и хондроцитами и улучшает костеобразование. Регулирует выживаемость, рост и «сервисное обслуживание» нейронов. После травм отвечает за репарацию и регенерацию нервных волокон. Модулирует синаптическую пластичность нейронов. Снижает количество фибриновых тромбов.
S 100 (кальпротектин) – принимает участие в воспалительных и регенеративных процессах за счет регуляции транспорта кальция.
Мусклин (Musclin, остеокрин) – повышает физическую выносливость и способствует биогенезу митохондрий. Регулирует рост костей (подавляет остеокластогенез и регулирует ремоделирование). Повышает захват глюкозы и синтез гликогена, защищает против потери мышечной ткани при раковых заболеваниях, контролирует кровяное давление и сократительную способность сосудов.
Остеонектин (SPARC) – регулирует ремоделирование скелетных мышц, подавляет адипогенез, влияет на системную толерантность к глюкозе. Регулирует костеобразование, подавляет рост опухолей. Участвует в накоплении коллагена и ремоделировании экстраклеточного матрикса. Отвечает за модуляцию экспрессии провоспалительных цитокинов, которые влияют на резистентность к инсулину.
Онкостатин-М – подавляет пролиферацию и индуцирует апоптоз раковых клеток (толстая кишка, грудь).
Металлопротеиназа– 2 (ММР – 2) – повышает клеточную пролиферацию и дифференцировку, поддерживает гомеостаз экстраклеточного матрикса. Обладает провоспалительными и противовоспалительными свойствами (зависит от цитокинового «фона»).
Фоллистатин подобный протеин 1 (FSTL – 1) (мышечный) – стимулирует захват глюкозы, влияет на функцию эндотелиальных клеток, повышает функционал бета–клеток, смягчает фиброз печени при функциональных повреждениях, поддерживает рост и развитие кардиомиоцитов. FSTL-1 отвечает за регенерацию сердечной мышцы за счет активации ангиогенных факторов.
Бета-аминоизомасляная кислота (BAIBA) – повышает митохондриальную функцию, усиливает противовоспалительные эффекты, снижает накопление жира, снижает фиброз почек, улучшает резистентность к инсулину и снижает воспаление в скелетных мышцах. Защищает остеоциты от воздействия активных форм кислорода (АФК) и предотвращает потерю костной массы при разгрузке нижних конечностей, уменьшает последствия глюкозно/липидных метаболических нарушений, вызывает «побурение» белого жира.
Катепсин В (CTSB) – усиливает выработку нейротрофического фактора головного мозга (BDNF) в гиппокампе и стимулирует нейрогенез, что благоприятно сказывается на когнитивных способностях; улучшает память, увеличивает нейрогенез в гиппокампе.
Wnt-3а (белок) (источник – костная ткань) – активирует миогенез и усиливает способность мышц к сокращению. Положительно регулирует костеобразование.
Склеростин (SOST) – отрицательно регулирует костеобразование через блокирование Wnts.
Белок 1, родственный Диккопфу (Dkk1) – отрицательно регулирует костеобразование через блокирование Wnts.
Фактор роста фибробластов-23 (FGF-23) (костный) отрицательно влияет на кардиомиоциты (мышцы сердца). Регулирует реабсорбцию фосфатов, поддерживая нормальное содержание минералов в костной ткани.
Фактор роста фибробластов-2 (FGF-2) – усиливает формирование кости и участвует в репарации костей.
Простагландин Е2 (PGE-2) (костный) – усиливает миогенез за счет стимулирования пролиферации миобластов и улучшает сократительную функцию мышц; регулирует рост костей.
Остеопрогерин (OPG) – подавляет активность остеокластов; продуцируется остеобластами под действием кальцитонина или эстрогенов.
Мионектин (CTRP 15) – повышает утилизацию глюкозы и стимулирует окисление жирных кислот
Апелин (Apelin) – контролирует кровяное давление и сократимость миокарда, снижает риск гипертрофии сердца, повышает усвоение глюкозы, улучшает когнитивные способности. Усиливает синтез ММР-1,3 и 9 и IL-1β. Снижает синтез коллагена II типа. Мышечный апелин улучшает митохондриальный биогенез, повышает синтез сократительных белков, стимулирует дифференцировку стволовых клеток по мышечному типу, препятствуя старческой саркопении.
Цилиарный нейротрофический фактор (CNTF) – нейротрофин (нейтрофин), обеспечивающий выживаемость нейронов, их развитие, пластичность синапсов и в качестве миокина (!) предотвращает оссификацию мышц; регулирует развитие периоста, при этом подавляет дифференцировку остеобластов и формирование кости, вызывая остеопороз.
Костный морфогенный протеин (BRINP3; FAMSC) – усиливает дифференцировку остеобластов (формирование кости).
Хемокин CXCL 10 – критический фактор ангиогенеза, связанного с физическими упражнениями.
Фактор роста эндотелия сосудов (VEGF) – PGC-1α зависимый – стимулирует рост новых капилляров и сосудов как реакция на рабочую гипоксию мышц.
L-Лактат – усиливает ангиогенез в головном мозге и улучшает мозговую функцию. Является посредником между метаболизмом жировой ткани и физической нагрузкой через стимуляцию синтеза трансформирующего фактора роста – бета 2. Регулирует иммунный ответ, принимает участие в репарации тканей и поддержании гомеостаза. Пиковый «постнагрузочный» лактат усиливает синтез коллагена II типа; при «перегрузке» системы лактатом при длительной интенсивной работе повышает синтез аггреканаз и металлопротеиназ, снижает синтез коллагена II типа (функция «контроль/ремоделирование»).
Хемокин CXCL12 – защищает от потери мышечной массы при кахексии, поддерживает рост мышц.
Дифференциальный фактор роста 15; макрофаг-подавляющий цитокин 1 (GDF -15) – противовоспалительный, снижает оксидативный стресс.
Активин А – принимает участие в регуляции «созревания» кости и хондрогенезе.
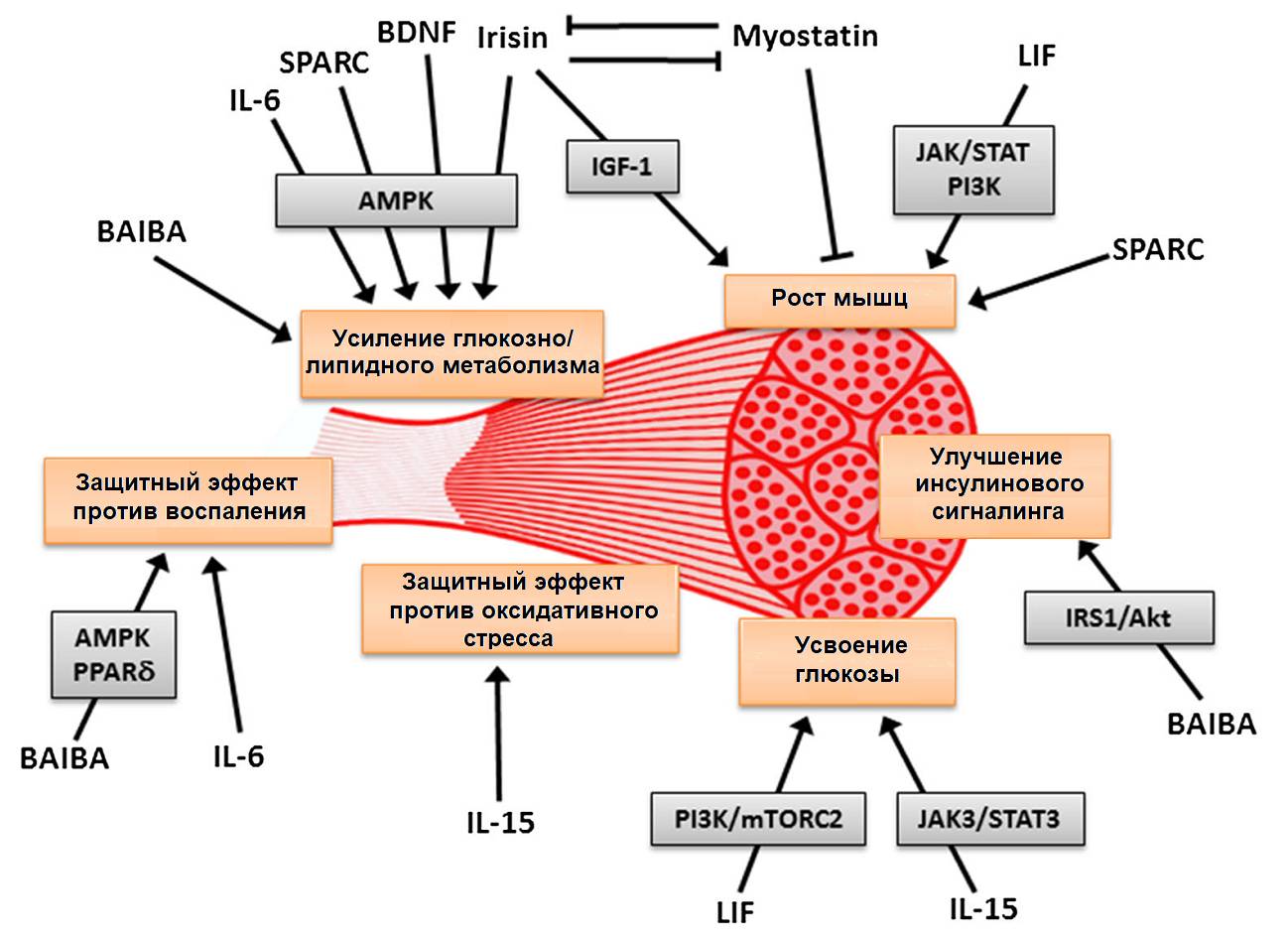
Основные миокины, их сигнальные пути и эффекты воздействия (Huh, 2018).
Итак. Физическая активность (упражнения) стимулирует высвобождение экзеркинов – цитокинов физической нагрузки. Сюда входят миокины, остеокины, кардиокины (здесь не упоминаются), гепатокины (не упоминаются), адипокины, цитокины и прочие «-кины».
Приведу примеры костно-мышечного перекрестного взаимодействия:
Нагрузка на кости скелета приводит к выбросу в кровь остеокальцина (OCN) и [костного] инсулиноподобного фактора роста-1 (IGF-1), которые стимулируют метаболизм мышц и вызывают их гипертрофию через усиление синтеза коактиватора-1α (PGC-1α). Связывание остеокальцина с андрогенным рецептором (GPCR6A) в мышцах усиливает адаптацию к физическим упражнениям и улучшает общую физическую форму. Остеокальцин усиливает синтез адипонектина [противовоспалительного] и витамина D. Остеокальцин способствует транслокации [перемещению] в ядро клетки активированного эритроидного фактора Nrf2 (антиоксидантная функция), что влияет на митохондриальный биогенез мышц. Остеокальцин стимулирует увеличение количества глюкозного транспортера 4 типа (GLUT4) на мембранах мышечных клеток, что усиливает усвоение глюкозы и гликолиз, таким образом повышая мышечную производительность.
Силовая нагрузка усиливает синтез остеокальцина, остеопрогерина и фактора роста фибробластов-23, при этом снижает синтез рецептора активатора ядерного фактора каппа би, его лиганда (RANK/RANKL) и склеростина. Остеопрогерин подавляет активацию рецептора RANK через связывание с RANKL [деактивация блокированием]. Лактат, который производится при интенсивных нагрузках, связывается со своим рецептором (Gpr81) на мембранах остеокластов и подавляет сигнальный путь ядерного фактора «каппа би» (NF-κB), предотвращая резорбцию кости. Также лактат стимулирует синтез трансформирующего фактора роста–2 бета, выступая как хондропротектор. Лактат стимулирует дифференцировку мезенхимальных стволовых клеток в остеобласты (костеобразующие). Излишек лактата после интенсивных физических нагрузок устраняется банальным растяжением мышц (самопроизвольным или принудительным) и успешно утилизируется тканями посредством кровотока.