



Станислав Дробышевский
Палеонтология антрополога. Том 3. Кайнозой
Некоторые отпечатки в Месселе столь качественны, что позволяют оценить цвет перьев по меланосомам: зимородок-щурка Eocoracias brachyptera был чудесной синей птичкой с чёрными крылышками и хвостиком.
В числе прочих пернатых эоцен подарил нам попугаев. Найти их в Индии – Vastanavis eocaena и V. cambayensis – не так удивительно, а вот в немецком Месселе – Psittacopes lepidus, Англии – Palaeopsittacus georgei – и Франции – Quercypsitta sudrei и Q. ivani – всё же неожиданно. Между прочим, эоценовые попугаи всё ещё имели довольно-таки непопугайный – тонкий и длинный – клюв, а их лапки были ещё полу-зигодактильными, то есть два пальца смотрели вперёд, один назад, а один поворачивался. С другими птицами география устраивает ещё более заковыристые выкрутасы: немецкие лягушкороты Masilopodargus longipes из Месселя родственны вайомингским Fluvioviridavis platyrhamphus из Грин Ривер, при том что современные живут от Южной Индии до Австралии и Тасмании.
* * *
Млекопитающие эоцена чудесны: они уже не столь однообразны, как палеоценовые, но ещё далеко не так похожи на современных, чтобы быть совсем уж привычными и банальными.
До конца эпохи протянули многобугорчатые, последними были североамериканские Ectypodus lovei, чьи нижние зубы достигли в своём роде апофеоза: огромный резец спереди, огромный гребнистый премоляр в середине и два бугристых моляра в конце. На границе олигоцена они вымерли из-за конкуренции с грызунами и приматами и появления новых хищников. В Аргентине коротко-широкомордые Groeberia minoprioi – родичи многобугорчатых из группы Gondwanatheria – выжили и дали потомков в последующие эпохи.
Сумчатые Marsupialia продолжали процветать в самых разных местах: многочисленные виды Peratherium в Европе и Африке (они дожили аж до миоцена), Asiadidelphis в Казахстане. Особое внимание обращает на себя Anatoliadelphys maasae – очень примитивное хищное сумчатое из Турции. Эта зверюшка весила 3–4 кг – как кошка или сумчатая куница; на последнюю анатолиядельфис был, видимо, похож и по способу передвижения – частично наземный, частично древесно-лазающий довольно бодрый хищник-падальщик. Судя по широким третьим премолярам, анатолиядельфисы любили грызть что-то очень твёрдое, видимо, кости. Из всех евразийских сумчатых это был самый крупный хищник. Впрочем, Турция в тот момент (и задолго до) была островом – этакой Австралией в миниатюре, так что тут могли реализоваться всякие странные экологические связи.
Гораздо более суровые хищники получились из сумчатых в Южной Америке, где сформировались Sparassodonta, или Borhyaenoidea, в лице первой группы Proborhyaenidae. Начав, как и все, с опоссума размером с куницу, они быстро превратились в двухметровых тридцатикилограммовых Callistoe vincei. Череп каллисто был хоть и узкий, но очень мощный, а главное – с огромными клыками на верхней и нижней челюстях, причем нижние были едва ли не больше верхних. Самое замечательное, что клыки не имели эмали и могли неограниченно или почти неограниченно расти, подобно резцам грызунов. Мощнейшие лапы с огромными когтями явно были приспособлены к рытью. Видимо, главной добычей каллисто были норные животные типа броненосцев – тогда и постояннорастущие клыки обретают особый смысл. Порой диггерский пыл каллисто зашкаливал: на типовом скелете локтевая и лучевая кости имеют следы заросшего перелома. Arminiheringia auceta, A. cultrata и A. contigua были массивнее – до 80 кг – и с несколько иными зубами: нижние клыки были растопырены, а заклыковые зубы туповаты, что выдаёт падальщика и всеядного зверя.
Как и прочие существа, сумчатые любили путешествовать: эоценовые южноамериканские опоссумы Zeusdelphys complicatus более родственны меловым североамериканским, а не местным, с которыми они, впрочем, конвергентно очень схожи. Видимо, это говорит о дополнительных миграциях.
Продолжались и прогулки по Антарктиде, где по острову Симор скакали Antarctodolops dailyi и A. mesetaense, представители специфического отряда Polydolopimorphia.
В палеоцене в Австралии млекопитающие неизвестны, но в эоцене этот недостаток был восполнен. Путь сумчатых из Южной Америки в Австралию прослеживается по зубам соневидных опоссумов Microbiotheria, которые те оставляли, двигаясь по дремучим антарктическим лесам, как заблудившиеся детишки из сказок, бросавшие хлебные крошки, чтобы найти обратный путь. Изолированные зубы подобных тварюшек найдены в среднеэоценовых отложениях всё того же западноантарктического острова Симор – Marambiotherium glacialis и Woodburnodon casei – и раннеэоценовых слоях фауны тингамарра в северо-восточной Австралии, где древнейшее сумчатое Djarthia murgonensis выглядит чрезвычайно генерализованно, но ближе всего именно к соневидным опоссумам. Также очень наглядно существование вида Chulpasia mattaueri в Перу и вида того же рода Ch. jimthorselli в раннеэоценовой Австралии; правда, определение австралийской находки вызывает сомнения, но как минимум сумчатость её несомненна.
Кстати, в последующем сумчатые переправились из Австралии в Индонезию, почти завершив удивительное кругосветное путешествие длиной в миллионы лет, ведь древнейшее сумчатое известно из Китая!
Работа над ошибками
В популярных книгах по биологии часто звучит, что Австралия – континент, заселённый только сумчатыми, дескать, из плацентарных сюда смогли долететь лишь летучие мыши да доплыть люди с собаками-динго. На самом же деле, в Австралии и на окружающих архипелагах живёт с полсотни эндемичных видов грызунов, попавших сюда совершенно независимо от людей.
Плацентарные эоцена демонстрируют наиширочайший спектр существ: от совершенно банальных до почти-фантастических.
Продвинутые цимолесты Pantolesta дали несколько разных линий. Одна из них – выдроподобная версия, представленная множеством видов североамериканских Palaeosinopa и Pantolestes, причём палеосинопы появились ещё в конце палеоцена в Северной Америке, а потом расселились и в Европу, где их потомки – Buxolestes piscator и B. minor найдены в разных местах, но лучшие отпечатки, конечно, в Месселе. Это были вытянутые приземистые зверьки почти метровой длины на коротких сильных гребных лапах с пальцами, как у бобра, с мощным уплощённым хвостом, развитыми спинными мышцами и мощными молярами для разжёвывания моллюсков и рыб. Внутри желудка на некоторых отпечатках сохранились останки рыб, так что образ жизни вырисовывается весьма очевидный.
Совсем другая линия – Ptolemaiida – не то часть пантолестов, не то самостоятельный отряд, не то часть афротериев, родственная трубкозубам (первоначально же они были описаны вообще как приматы). Они известны с границы эоцена и олигоцена египетского Файюма и Кении, а последние в Кении дотянули до миоцена. Лучше всего сохранился череп Ptolemaia lyonsi. Это были довольно крупные – с большую собаку – животные с плоской узкой вытянутой головой. Зубы птолемайид высококоронковые гипсодонтные, с тонкой эмалью, но с округлыми бугорками, как у приматов, а не складками эмали, как у копытных. Большие клыки сочетались с огромными третьими и четвёртыми премолярами и маленькими молярами. Челюсти хорошо двигались по вертикали, но плохо жевали. Не исключено, что такими зубами птолемайи разгрызали насекомых или моллюсков.
Среди насекомоядных в начале эоцена появляются первые достоверные ежи – китайские Changlelestes dissetiformis, а уже на границе с олигоценом – предки кротов – европейские Amphidozotherium cayluxi и Eotalpa anglica. Если вдуматься, и ежи с их иголками, и кроты с их лапами-лопатами и потерей зрения – крайне необычные твари, а кажутся привычными только потому, что мы знаем их с детства.
Среди прочих раннеэоценовых насекомоядных выделяется Batodonoides vanhouteni – при весе 0,93–1,82 г, в среднем 1,3 г, это было одно из самых маленьких млекопитающих всех времён! Очевидно, что такие микромасштабы батодоноидес мог иметь только в сверхтёплом климате и сверхобилии насекомых.
Как вы там, потомки?
Некоторые современные теплокровные приближаются к рекорду батодоноидеса, но всё же в полтора-два раза крупнее. Из млекопитающих это этрусская землеройка Suncus etruscus длиной 3–4,5 см (плюс хвост 3,5 см) и весом от 1,2 до 2,7 грамма, а также свиноносая летучая мышь Craseonycteris thonglongyai из Таиланда и Бирмы – 2,9–3,3 см и 1,5–3 г. Среди птиц свои рекордсмены: ямайская колибри карликовая пчёлка Mellisuga minima имеет длину с клювом 6–7 см, самцы весят 2–2,4 г, максимум 2,7 г, а самки весом 3,6 г несут яйца 1 см длины и 0,375 г весом; у кубинских колибри-пчёлок Mellisuga helenae самцы длиной 5,51 см и весом 1,6–1,95 г, самки – 6,12 см и 2,6 г, яйца длиной 6,35–11,4 мм.
Вообще, наземной и почвенной еды – червей, муравьёв, термитов, жуков, личинок, клубней, луковиц – в эоцене более чем хватало, так что не странно, что параллельно возникло несколько похожих линий ценителей подобного меню.
В Южной Америке таковыми стали неполнозубые Xenarthra. Первые известны из начала эпохи по щиткам панциря Riostegotherium и другим подобным из Бразилии и Аргентины, но толком про них ничего не известно. Впрочем, в том же раннем эоцене жил и хорошо сохранившийся броненосец Utaetus buccatus, уже вполне узнаваемый, но ещё сохранявший эмаль на редких колышковидных зубах, чего у более поздних и современных броненосцев уже нет.
Маленькая тонкость
Положение неполнозубых среди млекопитающих – великая загадка, слишком уж они специфичны. Существует три основные версии: в первой неполнозубые выделяются за пределы всех прочих плацентарных, которые тогда обозначаются как Epitheria; во второй афротерии всё же отстоят от других плацентарных больше, чем неполнозубые, в этом случае все, кроме афротериев и неполнозубых, объединяются в Exafroplacentalia (или Notolegia); в третьей неполнозубые с афротериями объединяются в Atlantogenata и противопоставляются Boreoeutheria.
Как вы там, потомки?
Неполнозубые не просто так получили своё название, у них действительно не всё в порядке с зубами. Ленивцы и броненосцы не имеют ни резцов, ни клыков (только у двупалых ленивцев хорошо развиты клыки), а заклыковые зубы у них очень примитивные – одинаковые, колышковидные, без корней и эмали. Примитивное ли это состояние или же вторичное – неочевидно. У муравьедов зубов вообще нет. Скуловые дуги нормальные у броненосцев, хитро преобразованы в странные незамкнутые закорючки у ленивцев, а у муравьедов вообще редуцированы. Из-за всего этого неполнозубые фактически не могут жевать, да и кусают так себе, а потому имеют специальный мускульный отдел желудка. Чрезвычайно специализированы лапы и позвонки. В частности, на лопатке есть дополнительная ость, клювовидный отросток лопатки (формирующийся как самостоятельный коракоид и поздно прирастающий к лопатке) сливается с плечевым, когти на пальцах огромные, а число пальцев может сокращаться до двух или трёх. Число шейных позвонков у ленивцев отличается от стандартных для млекопитающих семи – от пяти до девяти, причём меняется индивидуально, грудных рекордное число – до 24–25, на позвонках есть дополнительные суставные поверхности, а седалищная кость таза сочленяется с хвостовыми позвонками. Кожа склонна к образованию ороговевших пластинок, которые у ископаемых гигантских ленивцев были подкожной бронёй, у муравьедов имеются в хвосте, а у броненосцев развиваются в полноценный панцирь. Уникален язык муравьеда – больше полуметра длиной, крепящийся аж к грудине, до которой достают и слюнные железы. Всё это вместе взятое говорит о долгой независимой эволюции группы, видимо, уходящей корнями в мезозой.
Некоторые существа зависают между отрядами, даже несмотря на хорошую сохранность образцов. Eurotamandua joresi из Месселя известен по отпечатку целой тушки, на которой видно все детали, но всё равно специалисты спорят, относится ли он к муравьедам из отряда неполнозубых, к панголинам или к особой группе Afredentata в составе афротериев. Внешне евротамандуа действительно сильно похож на тамандуа – современного муравьеда из Южной и Центральной Америки: вытянутая в трубочку беззубая морда, короткие мощные лапы с огромными когтями, цепкий хвост. Многим кажется проблемой география, ведь муравьеды во все времена считались сугубо южноамериканской группой, но среди самых разных животных эоцена полно примеров географических загадок. Для других более показательно отсутствие у евротамандуа чешуи, а ведь роговая чешуя, покрывающая тело как шишку, это едва ли не самый яркий признак современных панголинов.
Гораздо надёжнее относится к панголинам Euromanis krebsi и Eomanis waldi всё из того же Месселя. Спина эоманиса уже точно была покрыта чешуёй, хотя ноги и хвост пока ещё не забронировались. Между прочим, благодаря идеальной сохранности содержимого желудка мы знаем, что эоманисы ели не только насекомых, но и растения.
Получается, что в Европе экологическая ниша расковыривателей муравейников и термитников была занята панголинами Pholidota. Тем более здорово, что они быстро пробрались и в Северную Америку, где по деревьям лазали цепкохвостые панголины Patriomanis americana. Вероятно, ещё позже они обосновались в Африке и Азии, где и живут до сих пор.
Непривычнее для нашего северного взгляда Palaeanodonta – родственники панголинов или, возможно, неполнозубых (при том, что сами панголины и неполнозубые друг другу сильно неродственны). Палеанодонты были коротконогими и очень вытянутыми зверьками в полметра – метр длиной (причём половину составлял хвост) с весьма странным низким продолговатым плоским черепом. Зубы североамериканских палеанодонтов прогрессивно уменьшались: если у Alocodontulum atopum на нижней челюсти имелись резцы, клыки, три премоляра и три достаточно развитых моляра, то у Metacheiromys marshi и M. dasypus на верхней челюсти имелся лишь один клык и один редуцированный заклыковый зуб, а на нижней – один резец, клык и пара крошечных однокорневых заклыковых зубов. Судя по мощным лапам со здоровенными широкими когтями, палеанодонты выкапывали себе пищу из земли, а судя по пропаже зубов, пища не требовала пережёвывания. Тем более странно, что клыки-то у палеанодонтов довольно большие; возможно, они больше использовались для внутривидовых демонстраций, чем для питания.
Работа над ошибками
Первое описание Metacheiromys было сделано по фрагментам скелета, в который по недоразумению были включены части от каких-то приматов. Ошибочно посчитали, что конечности метахейромиса похожи на руконожковые, за что он и получил свое название (Cheiromys – один из её старых синонимов) и был расценён как примат. Просто удивительно, насколько метахейромис в действительности не похож ни на приматов в целом, ни на руконожку в частности – и внешне, и по образу жизни!
Спасаясь от изменений климата конца эоцена, некоторые палеанодонты изобретали экзотические адаптации. Североамериканские Xenocranium pileorivale и Epoicotherium unicum стали подземными копателями – аналогами кротов и златокротов. Их головы стали короткими, с широкими вогнутыми лопатообразными носами, зубы упростились и превратились в редкие колышки, хотя клыки остались большими и толстыми. Слуховые части невероятно увеличились и превратились в широко расставленные в стороны пузыри, слуховые косточки утяжелились и увеличились, обеспечивая низкочастотный подземный слух. Для отгребания земли шейные позвонки с третьего по пятый срослись, плечевая кость превратилась в странную раскоряченную загогулину, в которой узнать плечевую кость – уже непростая задача; столь же искажёнными оказались и кости предплечья, а кисть стала загребущей когтистой лопатой-ковшом. Показательно, что в тех же слоях часты и кости амфисбены Spathorhynchus natronicus.
Родственный ксенокраниуму и эпойкотерию раннеэоценовый североамериканский Tubulodon выдвигался в качестве кандидата на предка трубкозубов, так как его зубы имеют дентиновые трубочки, но эта связь вряд ли оправданна, так как трубочки у него не той системы.
Травоядные эоцена весьма разнообразны. Крупнейшими были Pantodonta. В начале эпохи клыкастые тапироподобные Coryphodon eocaenus немножко уменьшились по сравнению с позднепалеоценовыми, но потом снова достигли размеров коровы и веса 700 кг у позднейшего вида C. lobatus; правда, судьбинушка у них не заладилась, и в середине эоцена корифодоны вымерли. Корифодоны жили в жарких тропических болотах, тем более примечательно обнаружение вида C. pisuqti на острове Элсмир, расположенном рядом с северной Гренландией. Эх, хорошо было в эоцене! На границе палеоцена и эоцена корифодоны проникли из Северной Америки в Монголию, Казахстан и Китай, а чуть позже – и в Европу. Последний представитель группы – Hypercoryphodon thomsoni – дожил до самого конца эоцена или даже олигоцена во Внутренней Монголии. Он достиг размера носорога. Череп гиперкорифодона выглядит средним между свиным и носорожьим, только с узкими скуловыми дугами. Колоритен и Eudinoceras mongoliensis со свиноподобной нижней челюстью и торчащими в стороны нижними клыками.
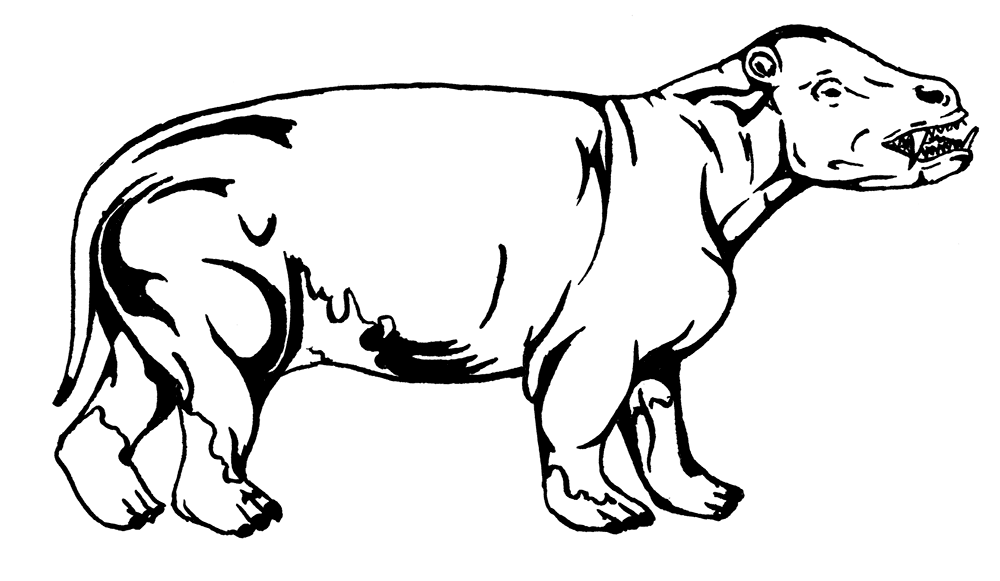
Coryphodon eocaenus
Конкуренты пантодонтов тоже не стояли на месте. Tillodontia тоже только в эоцене попали в Европу. В своём роде апофеозом развития группы стал Trogosus – крупный растительноядный зверь, отдалённо похожий на хвостатого медведя весом в полтораста килограмм. Тридцатисантиметровый череп с мощнейшими челюстями впечатляет, но, судя по огромным постояннорастущим резцам и стёртым молярам, трогозусы питались какой-то грубой растительной пищей типа корневищ.
Среди Taeniodonta самым крупным был Stylinodon mirus – весом в 80 кг, а его родич S. inexplicatus – вдвое меньше. Мордой стилинодоны были более чем импозантны: огромнейшая нижняя челюсть, которой позавидовал бы любой супермен, огромнейшие боковые резцы и особенно клыки, росшие на протяжении всей жизни, подобно грызунячьим резцам, огромнейшие когти на коренастых лапищах, причём передние были заметно сильнее задних. Видимо, эти монстры выкапывали своими граблями грубоволокнистые корни и клубни и грызли их своими зубищами.
Огромные грызущие звери были очень колоритны, но не могли противостоять новой группе – грызунам Rodentia. Отличный грызущий аппарат, многочисленность и быстрое размножение сделали грызунов сверхуспешными; неспроста нынче это самый многочисленный и по количеству видов, и по количеству особей отряд млекопитающих. Эоценовых грызунов уже очень много. Судя по костям лапок крайне примитивных грызунов Tamquammys robustus и T. wilsoni, двигались они примерно как крысы, может, чуть лучше лазая по ветвям. Особенно удались многочисленные виды белкоподобных Paramys, расселившиеся и по Северной Америке, и по Европе, и по Азии, а также чуть более поздних и более крупных североамериканских Ischyromys. Иширомису родственен и древесный цепкохвостый «дикобраз» Ailuravus macrurus из Месселя.
Очень показательна эволюция мозга грызунов, она шла ровно в тех же направлениях, что и эволюция мозга ранних приматов: мозжечок и особенно полушария конечного мозга увеличивались, а обонятельные доли уменьшались. На первых порах парамисы имели даже относительно больший размер головного мозга, чем синхронные ранние приматоморфы! Правда, процент развития новой коры конечного мозга у грызунов отставал, а темпы прогресса у наших предков были куда бодрее, так что мы вырвались вперёд и стали умничками-разумничками, а белки стали белками.
Как и приматы, первые грызуны освоили древесный образ жизни и питание растениями, но сделали это по-своему. Приматы научились далеко скакать, а грызуны предпочитают перелезать с ветки на ветку без длинных прыжков. Приматы налегли на сочные фрукты, а грызуны – на сухие семена и орехи. Также приматы оказались более-менее универсалами, тогда как грызуны очень специализированно приспосабливаются к каждой конкретной экологической нише с образованием новых видов. Те приматы, которые пытались пойти по грызунячьему пути – несколько линий плезиадаписовых с огромными резцами, – в итоге вымерли, так как, во-первых, не смогли развить столь совершенную жевательную мускулатуру, а во-вторых, вероятно, медленнее плодились.
В самом конце эоцена возникли архаичные бобры Agnotocastor и Anchitheriomys: с этого момента судьба водоёмов была предопределена; впрочем, многие потомки первых бобров стали наземными и даже норными, просто до наших дней дошли лишь любители плавания.
Прорыв грызуны совершили и в географическом смысле: ещё в середине эоцена, около 41 миллиона лет назад в Южной Америке появляются первые кавиоморфные грызуны Caviomorpha – всем известна морская свинка, а кто-то знает и шиншиллу, дегу и капибару. Первые представители – Cachiyacuy contamanensis, C. kummeli и Canaanimys maquiensis – найдены в Перу, а ведь ближайшие более-менее синхронные родственники жили в Африке, Южной и Восточной Азии, но не в Северной Америке, которая к тому же была отделена от Южной проливом. Стало быть, грызуны преодолели Атлантический океан напрямую! И стало быть, до Перу протошиншиллам надо было ещё добираться поперёк всего континента.
Как вы там, потомки?
Один из ближайших современных внеамериканских родственников кавиоморфных грызунов – Laonastes aenigmamus – неожиданно нашёлся в Лаосе (причём сначала на мясном рынке, потом в совиных погадках и лишь после целенаправленных поисков – вживе). Эта серая «крысобелка» оказалась достойной выделения в самостоятельное семейство Laonastidae или включения в семейство Diatomyidae, известное по нескольким ископаемым родам. Ясно, что заселение Южной Америки не шло из Юго-Восточной Азии через Тихий океан и даже не через Северную Америку, просто потомки неких африканских предков разбрелись в разные стороны: одни поплыли на запад, другие побежали на восток. В самой Африке тоже остались родичи – землекопы Bathyergidae и скальные крысы Petromus typicus.
Своим путём двинулись другие представители Mixodontia из семейства Mimotonidae – монгольские Gomphos elkema и киргизские Anatolmylus rozhdestuenskii: они и их родственники стоят в основании зайцеобразных Lagomorpha. В отличие от грызунов, они научились быстрее бегать, а также питаться корой и травой. По сравнению с грызунами, зайцеобразные стали заметно более универсальными: число их видов гораздо меньше, а экологическая пластичность некоторых видов – гораздо больше. Правда, грызунов такое количество и разнообразие, что любые масштабные обобщения в любом случае натыкаются на обилие исключений.
Маленькая тонкость
Существует два крайних полюса экологической пластичности – терпимости – устойчивости живых существ: стенобионты имеют очень жёсткие требования к среде и живут в узком диапазоне условий, а эврибионты – универсалы, способны жить где попало и есть что придётся. Отличные примеры стенобионтов – панды, коалы и муравьеды, а эврибионтов – волки и люди.
Широкое распространение грызунов, вместе с приматами, привело к сокращению числа, а потом и вымиранию многобугорчатых Multituberculata. Как минимум с середины эоцена грызуны начинают отвоёвывать ресурсы и у копытных; по крайней мере, грызунов постоянно становится всё больше, а копытных – относительно меньше.
В преддверии новых времён возникали и ещё более новаторские группы. Чрезвычайно успешными, без сомнения, оказались рукокрылые Chiroptera, ведь они в итоге стали вторым после грызунов по числу видов отрядом млекопитающих. Кто породил летучих мышей – пока неизвестно, видимо, это были какие-то палеоценовые насекомоядные типа цимолестов; прямые предки наверняка давно уже найдены, но таятся в обилии трудноразличимых палеоценовых зубов. Даже у стопроцентных среднеэоценовых летучих мышей зубы сложно отличить от землеройковых, что уж говорить о пращурах.
Древнейшая раннеэоценовая летучая мышь Australonycteris clarkae найдена в Австралии; очевидно, что туда надо было ещё долететь, так как в самой Австралии возникнуть было вроде как не из кого. При этом она ещё и не самая примитивная и даже – правда, сугубо предположительно – уже могла иметь способности к эхолокации. К сожалению, от австралониктериса сохранилось совсем немного. Зато практически идеально сохранились скелетики едва более поздних Onychonycteris finneyi из Вайоминга – самых примитивных летучих мышек. В частности, у онихониктериса сохранялсь коготочки на всех пяти пальцах ручки, не было эхолокации, зато был длинный хвостик. Однако все прочие летучемышьи особенности выражены практически полностью, крылья большие и полноценные, хотя некоторые аэродинамические нюансы и отличаются от современных. После летучих мышей становилось только больше и больше: Icaronycteris index, I. menui и I. sigei из Северной Америки, Франции и Индии, с пяток видов Archaeonycteris из Европы и Индии – и прочая, и прочая.
Маленькая тонкость
Рукокрылые – идеальный пример сверхбыстрой эволюции в начале группы, последующей остановки прогресса на большом таксономическом уровне после достижения оптимума и интенсивного видообразования во все времена. В палеоцене мы не видим летучих мышей ни в каком виде, в начале эоцена они возникают практически в готовом виде, а потом у них ничего принципиально не меняется, но виды с частными видоизменениями возникают в огромном количестве. Тут-то бы и закричать всяким креационистам о сотворении, но вот только такая модель эволюции многократно повторялась и до, и после в самых разных группах. То же самое было и с акулами, и с двоякодышащими рыбами, и с черепахами, и с птицами, и с бесчисленным количеством самых разных тварей, причём, чем специфичнее образ жизни и адаптации, тем лучше это выражено. Обрести машущий полёт трудно, для этого должны сойтись многие преадаптации в своеобразных условиях жизни. Зато в дальнейшем поменять рукокрылую специализацию почти невозможно: из летучемышьих крылышек и ножек, носа и уха уже не получишь ни ласты, ни хоботы. В принципе, человек со своей разумностью – из той же серии: возник один раз при странном стечении обстоятельств, достиг своего идеала и не знает, куда эволюционировать дальше.
Рукокрылые, наряду с грызунами, стали одними из главных наших конкурентов, только с другой стороны дерева: если протомыши не пускали приматов на землю и лучше грызли сухие семена, то летучие мыши не дали нам взлететь в воздух и вольготно гоняться там за комарами и бабочками. Осталось прыгать по кронам и искать плоды.
* * *
Траво- и листоядные в эоцене в условиях богатейшего развития растительности получили полный карт-бланш.
В Северной Америке и Азии расцвели Dinocerata, или Uintatheria. Твари удались на славу. Монгольские Gobiatherium бултыхались в болотах, рассекая тину огромным носовым гребнем с гигантскими ноздрями. Североамериканские Eobasileus cornutus и Uintatherium anceps (он же Dinoceras), а также китайские Uintatherium insperatus поражают своими метровыми башками: на носу, над глазами и на затылке возвышались огромные костяные шишки-рога, а из пасти торчали длинные изогнутые клыки, которым позавидовал бы любой саблезубый тигр. При этом в целом животные были похожи скорее на длинноногих носорогов и питались растениями. Думается, впечатляющие украшения были созданы половым отбором и использовались для турниров между самцами; саблезубость же типична для очень многих архаичных млекопитающих.
Маленькая тонкость
Показательно, что в других линиях копытных отростки на голове чаще возникали не как выросты черепа, а как производные кожи: у оленей они в итоге окостеневают, но площадь их соединения с черепом очень мала, у баранов это роговые образования поверх костяных колышков, у носорогов – утрамбованные волосы. При ударах рог и волосы пружинят, а колебания с них не передаются напрямую на кости мозговой коробки и мозги, а гасятся по пути. Уинтатерии же долбились непосредственно своими черепушками, так что импульс дзынькал прямиком по мозгам. Конечно, наверняка и у них поверх рога была какая-то оболочка, а мозгов было не так уж много, но счастью и процветанию такое устройство черепа явно не способствовало. Из современных копытных максимально к уинтатериевой системе близки жирафы, но они больше бодаются шеями, а не рогами.
На основе афротериев возникло сразу несколько линий крупных травоядных. Родственники эмбритопод Phenacolophidae – Zaisanolophus obscurus в Казахстане и Sanshuilophus zhaoi в Китае – тянутся ниточкой от скромных палеоценовых предков к многообещающим потомкам, а уже ранние Embrithopoda Hypsamasia seni в Турции и Palaeoamasia kansui, Crivadiatherium mackennai и C. iliescui в Румынии представляют развитие группы. Несмотря на принадлежность к афротериям, эта группа скорее продвигалась из Азии в Африку, а не наоборот. Среднеэоценовый намибийский Namatherium blackcrowense с его чрезвычайно широкими скуловыми дугами и высококоронковыми зубами, вероятно, был основателем арсиноитериев, с конца эпохи расселившихся по северной половине Чёрного Континента и достигших успеха уже в олигоцене.
В эоцене появились все главные группы современных афротериев. Достаточно архаично выглядят даманы Hyracoidea, например Seggeurius amourensis и Microhyrax lavocati из Алжира и Марокко и более поздние Bunohyrax matsumotoi из Алжира и Dimaitherium patnaiki из Египта. Видимо, с самого начала они были похожи на сурков. Примитивные даманы были мелкими, их мелкие резцы ещё не росли непрерывно, а теперь они большие и растут, как у грызунов; между резцами и клыками не было зазора-диастемы; клыки ещё иногда были двухкорневые, у современных же этих зубов иногда вовсе нет; лапки были снабжены коготочками, а не копытцами, а ножки были пальцеходящими, а не стопоходящими.
Как вы там, потомки?
Несмотря на неуклюжий вид и отсутствие хвоста, современные даманы очень здорово лазают по отвесным поверхностям. Для успешного причпокивания к камням и деревьям середины стоп могут подниматься, образуя присоску, а для пущего прилипательного эффекта выделяют резиноподобный пот. Кстати, «я на солнышке лежал, пятки кверху держал, вот они и запотели» – это про даманов, именно так они сбрасывают лишнее тепло, поскольку только подошвы могут потеть. Другая, довольно странная железа расположена на середине спины, она секретирует пахучее вещество при возбуждении; некоторые даманы при опасности оголяют её и поворачивают к хищнику. Показательно, что даманы до сих пор не стали окончательно растительноядными и, кроме травы, листьев и корневищ, регулярно едят насекомых. Рожают даманы от одного до трёх детёнышей, причём очень больших, с самого начала зрячих и пушистых, способных лазать по корягам и камням.
Гораздо более впечатляющие родственники даманов – хоботные Proboscidea. Начав с семнадцатикилограммовых «свинок», они опережающими темпами росли, отъедаясь на сочной болотной растительности Северной Африки. Уже в начале эоцена Daouitherium rebouli вымахал до 80–200 кг, Numidotherium koholense – до 250–300 кг и метра в холке, а на границе с олигоценом – двухметровый Arcanotherium savagei и Barytherium grave – уже до трёх метров в холке! Черепа первых слонов очень похожи на даманьи; головы быстро укорачивались и становились всё более лобастыми; носовое отверстие смещалось на лоб, свидетельствуя о постепенном развитии хобота. Нижние резцы превращались в лопатки на конце всё более узкой нижней челюсти, а верхние – в ещё короткие, но уже узнаваемые бивни.




