



Станислав Дробышевский
Палеонтология антрополога. Том 3. Кайнозой

Moeritherium lyonsi
Наверное, самый известные, хотя и не самые примитивные древние слоны – Moeritherium lyonsi и его родственники из Египта и Ливии. Надо сказать, череп меритерия, если игнорировать нижнюю челюсть и зубы, внешне (с точки зрения антрополога, конечно) довольно похож на череп огромной землеройки – от мелких палеоценовых предков не отвертишься. Зубная формула меритерия была ещё очень примитивной: три резца, клык и по три премоляра и моляра на верхней челюсти, два резца и по три премоляра и моляра на нижней. Короткие толстые бивни, сделанные из вторых резцов, вряд ли торчали изо рта. Сам же зверь был приземистым тапироподобным жителем болот. Слоновья форма мозга меритерия уже вполне узнаваема – кругленькая, укороченная и расширенная, но с огромными обонятельными луковицами.
Маленькая тонкость
Файюмский оазис – истинная родина слонов. В нескольких карьерах тут сохранилась богатейшая фауна эоцена и олигоцена – от черепах и крокодилов до китов, слонов и обезьян. Первые находки тут были сделаны Георгом Швейнфуртом в XIX веке, после ещё больше откопал Чарльз Эндрюс, а за ними подтянулись и другие исследователи, которые ныне являются классиками палеонтологии. Файюмские комплексы уникальны наличием и макро-, и микрофауны: чаще сохраняются останки либо больших, либо мелких зверей, а тут есть все. В итоге мы можем прослеживать изменения целой экосистемы на протяжении многих миллионов лет.
Файюм в эоцене и олигоцене представлял собой густо заросшие болота у берега моря, поэтому тут сохранилась и пресноводная, и солоноводная фауна. Удивительно найти посреди Сахары скелеты древнейших китов. Среди наземных млекопитающих огромный процент костей принадлежит именно афротериям, но есть и пришельцы с других континентов – сумчатые, креодонты, антракотерии, грызуны и приматы.
Родственная меритерию Saloumia gorodiskii известна лишь по одному обломку моляра, но важна тем, что жила в Сенегале. Это значит, что наши знания о североафриканских хоботных – лишь частичка реальной мозаики.
На границе эоцена и олигоцена слоны приобрели уже вполне узнаваемый облик: Palaeomastodon beadnelli (Proboscidea) и несколько других видов уже имели хорошо развитые верхние бивни (правда, не слишком длинные и загнутые вниз), утратили лишние резцы и клыки, обрели нормальный хобот и выросли до двух метров в холке. Нижняя челюсть приняла вид длинной лопаты с сошником из соединённых резцов для загребания болотной растительности.
Как вы там, потомки?
Слоны знамениты самыми разнообразными достоинствами – хоботом, ушами, бивнями, многосменными зубами, огромными мозгами и мощным интеллектом. Но мало кто вспоминает, что у слонов на ручках и ножках есть преполлекс и прехаллюкс – рудименты шестых пальцев (или нулевых, так как они расположены перед первым пальцем). Они развиты, конечно, не совсем полноценно, а в виде сесамоидов (косточек внутри связок), но почти настоящие. Слонам они нужны в качестве задней подпорки под тумбу кисти, когда слон переносит на неё вес тела. Болотная жизнь предков способствовала разлапистости слоновьих кистей и стоп, дабы не увязать в почве; благодаря этому первый палец не исчез, а это дало возможность развития предпервых сесамоидов для дополнительной опоры внутренней стороны кисти и стопы, а это, в свою очередь, обеспечило возможности для ещё большего увеличения размеров тела. У прочих копытных первый палец исчез слишком рано, отчего у них нет таких сесамоидов, что не даёт им стать по-настоящему гигантскими. Впрочем, у некоторых лошадей бывает рудимент большого пальца, так что для них не всё потеряно, надежда на появление многометровых сивок-бурок ещё теплится.
Между прочим, у нас нет никаких доказательств наличия предпервых пальцев у гигантских динозавров зауропод, но у них в силу исходной разлапистой рептильности сохранились другие большие пальцы, так что с опорой у них во все времена было хорошо, что и позволило вымахать до фантастических размеров.
Дополнительные недопальцы встречаются и у других зверей, причём по разным поводам: у панды дополнительный предпервый сесамоид служит для сгребания бамбука; у малой панды имеется такой же, но возникший независимо и конвергентно; у крота предпервый «палец» нужен для копания; у руконожки – для более успешного лазания по ветвям.
Слоны, появившись в болотах, смогли в итоге их покинуть, а вот их родственники сирены Sirenia, напротив, занырнули глубже, уплыв даже в моря. Несмотря на принадлежность к группе афротериев и теоретическую привязку к Африке, древнейшие сирены Prorastomus sirenoides ходили по раннеэоценовым берегам Ямайки. Мордой они были уже вполне узнаваемы, а вытянутые узкие челюсти были снабжены наиполнейшим набором зубов: три резца, клык, пять премоляров и три моляра. В середине эпохи двухметровый Pezosiren portelli на той же Ямайке по-прежнему сохранял хоть и коротенькие, но развитые ноги; тело же было очень длинным, уже вполне дирижабле-сиреновым; совершенно сиреновой была и голова, а зубная формула не поменялась. Хвост пьезосирена не имел увеличенных поперечных отростков, то есть был скорее как у выдры, а не веслообразный. Примитивнейший вид рода Protosiren – P. eothene – обнаруживается в Пакистане в море Тетис, а его многочисленная родня – в Северной Америке, Европе и египетском Файюме: Protosiren fraasi, более крупный и продвинутый P. smithae и другие. Такое универсальное распространение, очевидно, шло вдоль побережий мелководных морей; между прочим, современный дюгонь Dugong dugon тоже живёт вдоль всех берегов Индийского океана – от Меланезии и Австралии до Красного моря и Мадагаскара. Кстати, в том же среднем эоцене примитивные сирены известны и с Мадагаскара – Eotheroides lambondrano, а другие виды этого рода – из Индии и Египта. У испанского Sobrarbesiren cardieli всё ещё были четыре развитые ноги, но задние уже заметно уменьшились. Здорово, что в ряду многочисленных эоценовых и олигоценовых сирен можно проследить все промежуточные стадии уменьшения ног, превращения ручек в ласты, а также изменения морды и зубов.
Как вы там, потомки?
Современные сирены удивительны множеством особенностей. Они могут жить и в пресной, и в солёной воде и притом, в отличие от всех прочих крупных водных млекопитающих, остаются сугубо растительноядными; оттого у них усложнился желудок и, возможно, поэтому они имеют рекордно маленькое сердце. Задние ноги у них исчезли, хотя рудименты тазовых и бедренных костей всё ещё есть. Передние преобразованы в ласты, в которых, впрочем, вполне сохраняются чёткие сегменты, а у ламантинов намечены пальцы с копытцами. Несмотря на внешнюю редукцию, ламантины своими ручками могут подносить растения ко рту и прижимать детёнышей к груди. Хвост ламантинов имеет вид весла, а у дюгоней раздвоен, как у дельфина. У ламантинов шесть шейных позвонков, в отличие от почти всех млекопитающих, кроме ленивцев. Зубы у ламантинов не имеют внятного числа смен: в каждый момент времени функционируют от пяти до семи зубов, причём на место стёршихся передних сзади появляются новые и сдвигаются, как на конвейере. У дюгоней многочисленные предкоренные и коренные у молодых заменяются на всего пару коренных у взрослых. У ламантинов резцы вырастают, но рано исчезают, замещаясь на роговые пластины; у дюгоней резцы не только сохраняются всю жизнь, но и у самцов превращаются в короткие бивни. У стеллеровой коровы зубы вовсе исчезли, а вслед за ними – и она сама.
Другая линия афротериев пошла по насекомоядному пути: файюмский Dilambdogale gheerbranti стоит в основании тенреков, а Widanelfarasia bowni и W. rasmusseni, хоть и похожи на них же, но представляют, видимо, свою отдельную эволюционную линию. В то же время появляются и древнейшие прыгунчики, например Chambius kasserinensis, Nementchatherium senarhense и Herodotius pattersoni.
Как вы там, потомки?
Современные тенреки Tenrecoidea, или Tenrecomorpha, – очень своеобразные зверюшки. Большая их часть живёт на Мадагаскаре, но некоторые – в материковой Африке. Многие мадагаскарские тенреки конвергентны ежам: обыкновенный тенрек Tenrec ecaudatus выглядит как бесхвостый опоссум со спрятанными в шерсти иголками, полосатый тенрек Hemicentetes semispinosus – как чёрно-жёлтый недоёжик с мордочкой-трубочкой, редкими иголками и растрёпанным колючим хохлом на загривке, а ежиный тенрек Setifer setosus и малый тенрек Echinops telfari вообще внешне от ежей малоотличимы. Живут тенреки как на земле, в том числе в норах, так и на деревьях, питаются в основном насекомыми, но при случае не отказываются и от фруктов. Тенреки способны впадать в спячку в сухой сезон. Тенреки – рекордсмены среди плацентарных по плодовитости, самки могут иметь до 24 сосков и родить до 25 детёнышей! Это, кроме прочего, стало одним из тормозов на пути поумнения тенреков: на каждого детёныша приходится слишком уж мало мозга и родительского внимания, социальность у тенреков совсем слаба и ограничивается сбиванием в кучку в каком-нибудь дупле для согревания ночью.
Среди тенреков есть и масса других экологических вариантов: длиннохвостые тенреки Microgale больше всего похожи на землероек с чрезвычайно длинным хвостом, рисовые тенреки Oryzorictes – на помесь жирной землеройки с длиннохвостым кротом, болотные тенреки Limnogale mergulus – на водяную крысу с лысым хвостом и перепонками на задних лапках, они хорошо плавают и питаются водными растениями и рыбой. Выдровые землеройки Potamogale velox, Micropotamogale lamottei и M. ruwenzorii в Экваториальной Африке внешне и по образу жизни скорее похожи на выдр и питаются соответственно – раками, рыбой и лягушками.
Очевидно, мадагаскарская экосистема, столь бедная на зверей, взрастила спектр экологических вариантов из того, что было в распоряжении. Заодно, глядя на разнообразие тенреков, можно легко представить раннекайнозойскую эволюционную радиацию млекопитающих.
Прыгунчики Macroscelidea частично конвергентны тушканчикам: у них длинные тонюсенькие ножки, длинный тонюсенький хвостик и компактное пушистое тельце. Нос прыгунчиков превращён в смешной вечно-подвижный хоботок. А вот зубы их мало отличаются от зубов насекомоядных, в которых прыгунчики традиционно и включались. Образцом прыгунчиков может служить короткоухий прыгучик Macroscelides proboscideus, а чёрно-оранжевые хоботковые собачки Rhynchocyon даже очень красивы. По некоторым чертам строения зубов и черепа прыгунчики похожи на тупай, а иногда могут болеть малярией, что тоже отсылает к приматам. Между прочим, головной мозг прыгунчиков развит очень даже хорошо относительно размера тела и челюстей; прыгунчики очень бодры и постоянно находятся в движении, у них отлично развиты все органы чувств. В принципе, из такого первоисходника можно было бы получить кого-то и более разумного, тем более что и самка рождает обычно всего одного-двух детёнышей. Только вот специализация к прыганию и рытью подвела: хватательную и трудовую конечность из прыгунчиковой лапки не сделаешь! К тому же детёныши прыгунчиков рождаются практически готовыми – пушистыми, зрячими и способными бегать. Всё детство занимает пару месяцев, так что на обучение времени совсем нет; продолжительность же жизни, как, кстати, и у тенреков, всего год-полтора.
В то время как афротерии превращались в тварей от прыгунчиков до слонов, прочие плацентарные – бореэвтерии – эволюционировали по-своему. На протяжении всего эоцена продолжали бытовать поздние кондиляртры Condylarthra типа Hyopsodus и Phenacodus, похожие на помесь куницы с карликовым оленьком. Некоторые из них ещё могли лазать по деревьям, некоторые рыли норы, но всё больше и больше переходили к бегу по кустам и растительноядности. Показательно примитивное строение их мозга: новая кора была слабо распространена вниз и не имела борозд с извилинами, обонятельные луковицы были огромными, а мозжечок – маленьким.
Работа над ошибками
Удивительно обнаружение Tingamarra porterorum – кондиляртры из раннего эоцена Австралии. Пусть это лишь один-единственный моляр, но чрезвычайно важный, так как показывает возможность миграции плацентарных зверей из Южной Америки в Австралию. Чисто гипотетически Австралия могла быть заселена кондиляртрами, из которых могли бы возникнуть самые разнообразные оригинальные существа. Однако некоторые палеонтологи считают отложения с зубом тингамарры позднеэоценовыми или даже позднеолигоценовыми, а зуб – принадлежащим сумчатому типа примитивного опоссума. Тогда вся красивая гипотеза осыпается. Возможно, новые находки в будущем лучше прояснят загадку тингамарры.
Южноамериканские копытные эволюционировали своими путями. Особенно странно выглядела аргентинская Thomashuxleya externa из группы Notoungulata, одна из первых представительниц Toxodontia. Это было животное весом 180 кг с мордой клыкастой собаки-дегенератки, на крепких лапах, с маленьким хвостиком. Судя по зубам и челюстям, питалась она, видимо, растениями, хотя по внешнему виду это было неочевидно. Другие нотоунгуляты были странны по-своему: у Notostylops зубная формула могла меняться от 3143/3143 до 2033/2033. Описано множество видов нотостилопсов, но есть мнение, что дело не в систематике, а в величайшей индивидуальной изменчивости. В целом эти зверюшки были очень усреднёнными, генерализованными: собакоподобные твари размером с кролика, экологически что-то вроде всеядных енотов, но с резцами, как у грызунов.
Маленькая тонкость
Зубная формула отражает число зубов в зубном ряду верхней и, через дробь, нижней челюсти; зубы считаются только на одной стороне. Первой цифрой идут резцы, потом клыки, затем премоляры и последними – моляры.
Нотоунгуляты Pseudhyrax eutrachytheroides и P. strangulatus были аналогами даманов.
Работа над ошибками
Маленький нотоунгулят Notopithecus adapinus первоначально был описан как примат, однако в реальности был копытным, похожим на длинноногую мордастую крысу, неплохо лазавшую по деревьям.
В эоцене Аргентины появились также первые представители отряда Litopterna (или Notopterna) – Amilnedwardsia brevicula и Indalecia grandensis, родственные знаменитым и гораздо более поздним макраухениям.
Доживали свой век и последние представители Xenungulata – Etayoa bacatensis в Колумбии размером с собаку и более крупная Notoetayoa gargantuai из Аргентины.
Кроме Австралии, Африки и Южной Америки существовал и ещё один материк-остров – Индия, плававшая в эоцене посреди моря Тетис. Тут развились свои аналоги хоботных и американских копытных – отряд Anthracobunia, например, многочисленные виды Anthracobune и Cambaytherium. Антракобуниды настолько похожи на первых хоботных, что некоторые авторы считают их предками слонов. Другие, впрочем, определяют их лишь как семейство Anthracobunidae в пределах непарнокопытных. Как и большинство эоценовых растительноядных, антракобуниды внешне больше всего напоминали тапиров, но имели довольно стройные ноги и длинный тонкий хвост. Интересно, что, несмотря на изоляцию, многие другие животные эоценовых фаун Индии родственны европейским, африканским и, в меньшей степени, азиатским. Однако антракобуниды – эндемики Индии, они так и не смогли переселиться куда-то ещё и в последующем вымерли, видимо, не выдержав конкуренции с привычными нам копытными.
* * *
Североамериканские и евразийские копытные пошли двумя основными путями – непарно- и парнокопытным.
Маленькая тонкость
Различаются непарно- и парнокопытные, конечно, не только парностью пальцев на ножках, хотя этот признак был важен и надёжен уже в эоцене. Две эти группы сделали разные ставки на развитие жевательного аппарата и пищеварительного тракта: непарнокопытные стали гораздо эффективнее жевать первично и переваривать кишечником, а парнокопытные – жевать вторично и переваривать желудком. Проще говоря: лошадка сначала старательно жуёт, потом немножко переваривает желудком, а после долго – кишечником, а коровка – сначала глотает не жуя, потом частично переваривает желудком, потом отрыгивает и тщательно пережёвывает, потом снова глотает и снова переваривает желудком, а потом ещё и кишечником. В итоге у непарнокопытных однокамерный желудок и очень длинный и важный кишечник, а у парнокопытных – четырёхкамерный желудок и сравнительно менее значимый кишечник.
Эоценовые непарнокопытные быстро взяли курс на увеличение размеров и были склонны давать гигантские формы, тогда как парнокопытные чаще оставались мелочью.
Непарнокопытные Perissodactyla начали свой подъём к успеху. Первые представители – Palaeotheriidae в лице английского Hyracotherium leporinum, немецкого Propalaeotherium parvulum и французского Palaeotherium magnum, что логично, мало отличались от кондиляртр: размером с большую кошку или собаку обитатели густых зарослей, питающиеся листьями кустарников и плодами. На передних ногах у них было четыре пальца, а на задних – уже всего три; на низких молярах – шесть тупых бугорков без всяких режущих гребней. В дальнейшем они пошли несколькими альтернативными путями.
Маленькая тонкость
Иногда из палеотериев выделяется семейство Pachynolophidae, представленное, например, Pachynolophus и Anchilophus. Эти звери были доминирующими непарнокопытными среднего эоцена Европы, но к концу эпохи очень резко сократились в числе и к началу олигоцена полностью вымерли.
Многие ранние непарнокопытные по славной эоценовой традиции стали тапироподобными жителями болот. Группа тапиров в виде нескольких архаичных семейств возникла в Северной Америке. Это и древнейшие мелкие Heptodon, и стройные Helaletes nanus. Довольно быстро они попали в Азию, где известны от Монголии до Казахстана, например Desmatotherium. Каким-то образом они добрались и до острова Индии – Vastanolophus holbrooki и Cambaylophus vastanensis. Первобытные тапиры обычно были довольно мелкими и ещё не имели такого хобота, как современные, но принципиально схожи со своими потомками. Между прочим, несмотря на название отряда, тапиры по-прежнему имеют четыре пальца на передних ногах, но три – на задних.
Особняком стоят азиатские эоценовые тапиры Lophialetes expeditus и Schlosseria magister, имевшие по три пальца и на стопе, и на кисти. Лофиалетесы вообще обогнали своё время, благо обгонять они умели: в эпоху господства толстеньких пожирателей околоводной растительности лофиалетесы были быстробегающими стройными газелеподобными длинношеими потребителями высоко расположенных листьев открывающихся степей. Однако, несмотря на прогрессивность, лофиалетесы вымерли, не выдержав конкуренции с парнокопытными, а более архаичные формы тапиров сохранились.
Тапироподобны были и первые носороги – Yimengia, Triplopus, Hyrachyus, Lophiodon, Amynodon, Metamynodon и прочие, относящиеся к нескольким архаичным семействам. Многие из этих родов имели крайне широкое распространение – в Северной Америке, Европе и Азии. Amynodon известен также из Японии, а Hyrachyus – даже на Ямайке! Большинство из них были приземистыми бочкообразными коротконогими жителями болот и прибрежных зарослей и питались ветками и листьями кустарников. В отличие от тапиров, носороги сделали ставку на режущие свойства зубов, высококоронковость, большие размеры тела и, как это ни противоречит предыдущему пункту, – на способности к бегу. У некоторых архаичных носорогов сохранялись нормальные резцы и довольно большие клыки; на ногах часто по-прежнему было четыре пальца спереди и три – сзади; степные лошадеподобные Hyracodon и предки индрикотериев Forstercooperia уже сократили число пальцев на кисти до трёх. Между прочим, китайские среднеэоценовые индрикотерии Juxia приобрели свой классический облик, только размером были скромнее своих знаменитых олигоценовых потомков. Судя по отступанию носовых костей назад, у продвинутых Cadurcodon имелся небольшой хобот; эти болотные животные максимально походили на тапиров, да к тому же с большими вертикальными бивнями. Вот чего не было у первых носорогов, так это рогов! Не было их даже у первых настоящих носорогов семейства Rhinocerotidae – среднеэоценовых североамериканских Uintaceras radinscyi.
Вот у кого рога развились, так это у Brontotheriidae, большая часть которых известна из Северной Америки; между прочим, филогенетически бронтотерии родственнее лошадям, а не носорогам. Самые первые мелкие Eotitanops ещё имели простенькую морду, чуть более поздние Palaeosyops размером с корову и ещё более мощные Manteoceras уже обзавелись вздутием на носу, а Protitanops curryi, Protitan имели хотя бы и невысокие, зато раздвоенные рога на носу. Бронтотерии, начав, как и все, с размера карликовой свинки, быстро выросли в огромных чудищ до двух с половиной метров в холке, став самыми большими и массивными животными своего времени. Огромные остистые отростки первых грудных позвонков были нужны для крепления мощнейшей мускулатуры для поддержания тяжеленной головы, тому же соответствовали и толстые ноги. В итоге вид бронтотериев был куда как харизматичный. Самый широко-разрекламированный бронтотерий – Megacerops (он же Brontotherium и Brontops), хотя пара его тупых округлых рогов на оригинальных черепах выглядит далеко не так впечатляюще, как на реконструкциях. У самцов рога были больше, чем у самок, что уже намекает на половой отбор и брачные турниры. Как и в случае с уинтатериями, бодание костными рогами должно было создавать сложности для сохранности мозга; впрочем, миллионы лет бронтотериям это проблемой не казалось. Известен скелет бронтотерия с результатом неудачного брачного турнира – сломанным и заросшим с костяной мозолью ребром на правом боку.

Megacerops
Монгольские Embolotherium пошли куда дальше: их носовой рог стал широченной раздвоенной лопатой, причём одинаково большой и у самцов, и у самок. Борозда на передней стороне шла от носовой полости и, возможно, служила для размещения резонатора: монгольские болота оглашались трубными кликами эмболотериев. Судя по румынскому Brachydiastematherium transylvanicum, подобные монстры жили и в Европе.
Эволюция лошадей – классика палеонтологии, каждый видел в школьном учебнике схему их эволюции, составленную В. О. Ковалевским. А начинается она с североамериканского Eohippus angustidens – первого представителя семейства Equidae. Эогиппус очень мало отличался от гиракотерия (а иногда они даже объединяются): всего 30 см в холке, со среднюю собачонку. Коронки его зубов были низкими, на ногах, как у всех примитивных непарнокопытных, по четыре пальца на передних и по три – на задних, из чего следует, что жил он в густых зарослях, бегал по влажной почве и питался листвой и тонкими ветками.
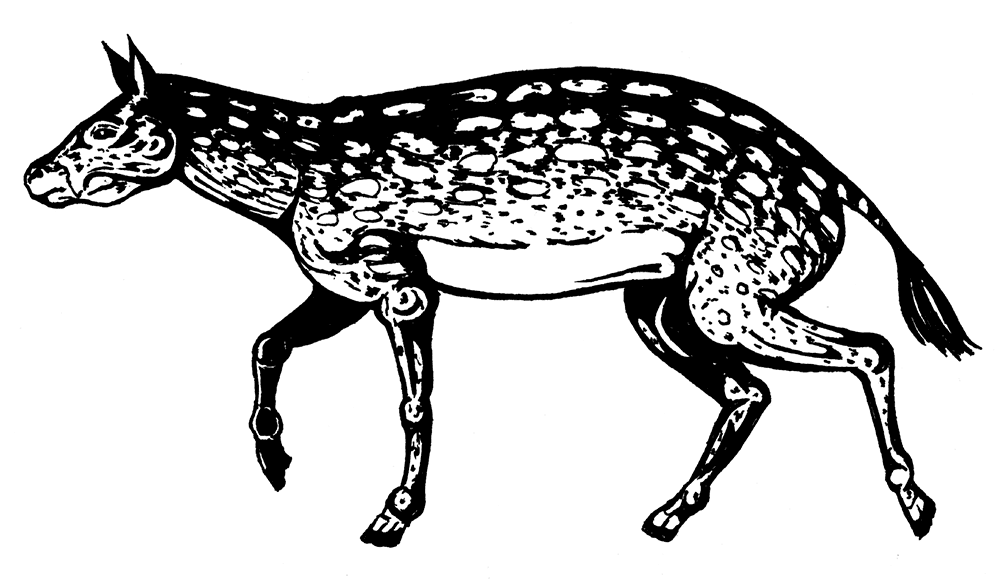
Eohippus angustidens
По мере изменения условий жизни исходная «кондиляртность» уменьшалась, а «лошадность» усиливалась, причём в нескольких эволюционных линиях независимо и немножко по-разному. Лошадиная ветвь представлена, например, последовательными Orohippus и Epihippus. У них всё ещё имелись по четыре пальца спереди и три сзади, а локтевая и лучевая кости предплечья оставались независимыми в память о роющих предках; по-прежнему низкокоронковые зубы помаленьку обзаводились гребнями для разрезания листьев; размеры тела помаленьку росли.
Маленькая тонкость
В. О. Ковалевский на основе тщательнейшего изучения эволюции конечностей и зубов ископаемых лошадей и энтелодонтов пришёл к выводу, что, во-первых, разные части тела эволюционируют с неравными скоростями и, во-вторых, в случае обгона одной системы другой общая итоговая приспособленность может оказаться ниже, чем у конкурентов, у которых признаки менялись медленнее, но равномернее. В. О. Ковалевский разделил понятия инадаптивной эволюции – неравномерной и не вполне согласованной, накладывающей ограничения на последующие прогрессивные изменения, и адаптивной – более соразмерной, без крайностей и заскоков. Кроме того, В. О. Ковалевский первым в палеонтологии развил и последовательно применял морфофункциональный анализ – оценку эволюционных изменений с позиции их приспособительного смысла в данной конкретной экологической среде. Сейчас такой подход кажется само собой разумеющимся, но в XIX веке это был мощнейший прорыв; стоит ведь помнить, что эволюционная теория только-только была опубликована, а палеонтология возникала на глазах. Да и сейчас во многих палеонтологических трудах занудное перечисление морфологических отличий следует без малейшей попытки осознать их смысл.
В эоцене среди лошадеобразных выделились Eomoropidae, например Paleomoropus jepseni и Eomoropus из Восточной Азии и Северной Америки. Эти листоядные звери размером с овцу имели довольно крупные клыки в пасти и когти на пальцах и могут быть предками более поздних халикотериев. Некоторые подобные животные, например европейский Lophiaspis, иногда уже напрямую относятся к халикотериям; впрочем, эоморопиды могли быть и параллельно развивавшейся группой.
Примитивные парнокопытные Artiodactyla в раннем эоцене эволюционировали несколько медленнее, чем непарнокопытные, зато раскачались к концу эпохи. Первые представители – Dichobunidae в лице многих видов раннеэоценовых Diacodexis, среднеэоценовых Homacodon и прочих других – широчайше распространились по Европе, Северной Америке и Азии. Внешне они выглядели как карликовые оленьки с длинными тонкими хвостиками весом два-три килограмма; несмотря на принадлежность к парнокопытным, они имели ключицы и по пять пальцев с копытцами на каждой ножке. Подобны им, разве что крупнее и с короткими хвостиками, были и североамериканские Hypertragulidae. Удивительна рассинхронизированность эволюции передних и задних конечностей: у Hypertragulus всё ещё пятипалая когтистая кисть сочеталась с функционально двупалой стопой (сохранялись рудиментарные фаланги ещё одного пальца стопы, но совсем крошечные), а у Leptomeryx – четырёхпалая ручка с полностью двупалой ножкой. Показательны изменения зубов: верхние резцы уже успели исчезнуть, но у гипертрагулюса имелись довольно крупные клыки и острые премоляры, а нижние резцы были почти вертикальны, тогда как у лептомерикса клыки уменьшились, премоляры почти перестали отличаться от моляров, а нижние резцы направились горизонтально вперёд. Несмотря на примитивность, гипертрагулюсы дожили до миоцена.
Азиатские Archaeomeryx optatus и его родственники размером с зайца были и внешне похожи на зайцев: попрыгунчики с длинными задними и короткими передними ножками-шпильками, развитыми верхними резцами, но ещё и с клыками, и с длинным хвостом. Очень быстро, уже к концу эоцена, подобные существа доэволюционировали до семейства Tragulidae, примерами чего могут быть Archaeotragulus krabiensis и Krabitherium waileki из Таиланда.
Как вы там, потомки?
Современные карликовые оленьки Tragulidae – азиатские Tragulus и Moschiola и африканские Hyemoschus aquaticus – просто напрашиваются на то, чтобы стать объектом загадок: саблезубый зверь, отлично плавает и даже ходит по дну, где ловит рыбу и раков, лазает по деревьям и лианам, день проводит в дупле, ночами выходит на поиски листьев, плодов, крыс, ящериц и всяческой падали. Кроме огромных клыков у самцов, оленьки обладают невероятно длинным языком и трёхкамерным желудком, а никаких намёков на рога у них нет. Весят они от полкило до восьми килограмм, африканские гиганты – до пятнадцати. По итогу, оленьки – едва ли не самые примитивные из нынешних парнокопытных, в их лице можно видеть осколок эоценового мира влажных тропических джунглей.
Саблезубость присуща и некоторым другим парнокопытным, например кабарге Moschus moschiferus, водяному оленю Hydropotes inermis и мунтжаку Muntiacus muntjak, причём у последнего клыки сочетаются с рогами.
Успешной и многочисленной группой эоценовых парнокопытных были Anthracotheriidae, например, древнейший мелкий Elomeryx и поздний двухсоткилограммовый Bothriodon (он же Ancodus); появились они, видимо, в Азии, потом оказались в Европе, а оттуда добрались и до Северной Америки в одну сторону и Африки – в другую. Особенно преуспели многочисленные виды Anthracotherium, дожившие от среднего эоцена аж до миоцена. Эти звери выглядели как приземистые тонконогие свино-тапиро-бегемотики с вытянутыми мордами, и ведь вправду они оказались предками гиппопотамов, а по менее вероятной версии – родственниками свиней. Набор зубов был практически полным, на передних лапках имелось по четыре пальца, а на задних – по пять. Такое строение оказалось практически универсальным для жизни в заросших болотах. Впрочем, антракотерии ещё не были слишком специализированы и могли уклоняться в разные стороны: некрупный бирманский Siamotherium pondaungensis бегал по открытым лесам и был вполне всеядным, не слишком уважая листья. Видимо, подобные антракотерии или очень родственные животные стали основой и для монстрических энтелодонов, и, возможно, для китов.

Anthracotherium
Как вы там, потомки?
Современные копытные рождаются практически готовыми, работоспособными, иногда они даже почти сразу пытаются щипать траву, хотя, конечно, довольно долго ещё кормятся молоком. Повышенная самостоятельность резко повышает выживаемость, даже при небольшом числе детёнышей – у копытных обычно один на одни роды. Получается, что, с одной стороны, это высшая степень доделанности и прогресса на момент рождения среди млекопитающих, а с другой – парадоксальным образом копытные вернулись в идеал рептилий, у которых из яйца вылупляется уже готовая ящерка, только маленькая. По сравнению с примитивными млекопитающими, то есть нами, такая стратегия вроде как и продвинутая, но диалектически становится тормозом на пути развития разума, ведь если детёныш обеспечен с рождения всеми необходимыми программами поведения, то и обучение уже не нужно. Оттого мы – архаичны по сравнению с овцами, но мы – люди, а они – овцы.
Во второй половине эоцена часть парнокопытных Северной Америки стала увеличиваться в размере, превратившись в двухметровых кабаноподобных тварей Achaenodon – самых крупных парнокопытных в своих фаунах. Они были либо первыми и уже впечатляющими Entelodontidae, либо, вероятнее, гигантскими представителями Helohyidae. Ахенодоны сохранили весьма архаичную зубную систему, в том числе огромные толстые клыки, острые премоляры и бунодонтные – низкокоронковые бугорчатые – моляры, а их короткие массивные головы были чем-то средним между кондиляртровыми и свиными. Вероятно, эти монстры были не сугубо растительноядными, а хотя бы частично занимали нишу крупных падальщиков и могли грызть кости. Несколько позже их сместили с пьедестала Brachyhyops и Archaeotherium – уже несомненные представители энтелодонтов, расселившиеся и по Северной Америке, и по Азии, и по Европе, успешно перешедшие в олигоцен и давшие там великолепное потомство. Длинные челюсти археотериев были усажены примитивными зубами, в том числе огромными клыками, и снабжены крупными выростами для мощнейших жевательных мышц, о чём явно свидетельствуют и широкие скуловые дуги. Как и ахенодоны, археотерии, кроме растений, пожирали падаль; этому же способствовало и отличное обоняние, о чём мы можем судить по увеличенным обонятельным луковицам. Тяжеленная голова поддерживалась сильными мышцами, крепящимися на длинные остистые отростки грудных позвонков, отчего археотерии имели ссутуленный вид. Ноги, несмотря на сравнительно большую длину, не были приспособлены к быстрому бегу, а кисть и стопа адаптировались к ходьбе по влажному грунту. Эти твари стали конкурентами не только и столько растительноядным, сколько мезонихиям и креодонтам.




