



Орис Орис
Ииссиидиология. Основы. Том 5
Глава 3. Нейрофибриллы как связующая функциональная основа между Инфо-Творцами, «субтеррансивной ОДС» и биохимической деятельностью био-Творцов ЦНС
5.141. На функциях клауструмов мы позже ещё остановимся, а вот о функциях нейронных микротрубочек (нейрофибрилл) следует уже сейчас поговорить более основательно и детально, так как без глубокого понимания их роли в деятельности био-Творцов головного мозга вам сложно будет понять не только важность данной фазы онтогенеза, но и сам процесс последовательного и избирательного преобразования информационного потенциала ОДС и ФЛК (через АСТТМАЙ-РАА-А и виваксы) сначала в электрические и биохимические реализации био-Творцов белково-нуклеинового комплекса, а затем – в наши с вами психоментальные и физические реакции.
5.142. Напомню, что мозг представляет собой сложнейшую нейронную сеть, при этом сама нервная система образована как бы из двух условных частей, одна из которых генетически обусловлена (изначально получена от родителей в виде ядерной ДНК, находящейся в теле клетки), а другая – в виде отростков нейронов, соединённых друг с другом через синапсы, – постоянно изменяется и развивается в процессе непрерывного осуществления Фокусной Динамикой «личности» своего ротационного Цикла (жизнедеятельности индивидуума). Каждый нейрон (нервная клетка), в ядре которого содержится генетическая Информация, имеет один длинный (иногда – до метра) выходной отросток – аксон, покрытый миелиновой оболочкой, и от 1000 до 20 000 дендритов – коротких входных ветвящихся отростков, воспринимающих сигналы от других нейронов, рецепторных клеток или непосредственно от внешних раздражителей.
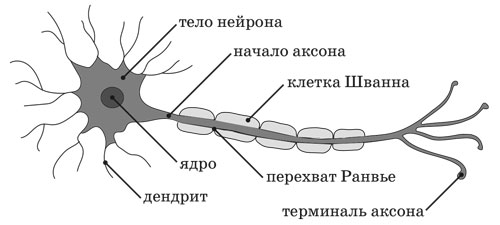
Рисунок 5.0142. Схема строения нейрона
5.143. Взаимосвязи между био-Творцами аксонов одних нейронов и био-Творцами дендритов других нейронов осуществляются через синапсы, чья работа – при координирующем участии био-Творцов коры головного мозга – обусловливает качественность нашей долговременной памяти, так как эффективность выполнения ими функций по передаче и приёму Энерго-Информации зависит от частоты их использования: при частом задействовании синтезируются новые нейромедиаторы, которые активизируют новые – более стабильные и эффективные – синаптические цепочки, а старые синапсы при этом прекращают свою деятельность. Изменение же активности самих синапсов (в результате преобразования Форм Самосознаний поступающих в них нейромедиаторов) отражается на свойствах нашей кратковременной памяти.
5.144. Ещё вы должны знать, что микротрубочки характерны не только для нейронов, но также являются важнейшей структурной единицей цитоскелета каждой клетки биологического организма, «сфероидально» распространяясь от центросомы (ядра клетки) ко всем участкам цитоплазматической мембраны. Они играют существенную роль в поддержании формы клетки и в транспортировке её органелл (включая митохондрии), а также во время деления клеток (через структуры центросомы) обеспечивают равномерное распределение генетического материала хромосом (геномной Информации) по дочерним клеткам. Поэтому био-Творцы нейрофибрилл всех нервных клеток (как ЦНС, так и периферической нервной системы) функционально очень тесно взаимосвязаны (через дендриты и аксоны) друг с другом и с реализационной динамикой био-Творцов микротрубочек всех клеток нашего биологического организма.
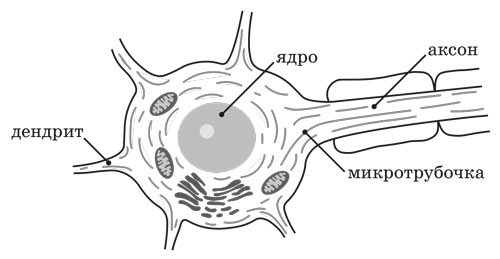
Рисунок 5.0144. Схема расположения нейронных микротрубочек
5.145. Каждая нейрофибрилла представляет собой своеобразный материальный носитель, специально «оборудованный» геномными Формо-Творцами для волнового приёма СФУУРММ-Форм какого-то каузального «Канала» от био-Творцов клауструмов и ретикулярной формации, с тем чтобы, параллельно химически закодировав данную Информацию в фокусные Конфигурации качественно соответствующих ей белковых носителей, тут же транслировать эти, заключённые в Формы Самосознаний трансмиттеров, СФУУРММ-Формы био-Творцам коры головного мозга, подкорки и далее – через всё множество цитоскелетных микротрубочек – био-Творцам каждой клетки организма. Всё это био-Творцы нейрофибрилл осуществляют под эгидой Творцов-интерпретаторов генома, деятельность которых, в свою очередь, нутационно корректируется Творцами-регуляторами ДНК, чьи ФС в данном диапазоне эксгиберации представляют реализационные Интересы ККР ЛЛУУ-ВВУ-Сущности.
5.146. Функциональные структуры каждой микротрубочки резонационно настроены на узкий спектр реализации Формо-Творцов какого-то одного каузального «Канала», нутационную динамику которого её био-Творцы (благодаря наличию тринадцати глобул молекул белка тубулина) дифференцируют внутри себя ещё на двенадцать, качественно отличающихся друг от друга, узкоспецифических реализационных подуровней (энергоинформационные параметры каждой из 12 глобул соответствуют вибрациям одного ДУУ-ЛЛИ-подуровня какого-то ИИССИИДИ-Центра; био-Творцы одной – тринадцатой – глобулы выполняют объединяюще-координирующие функции). В связи с наличием такой узкоспектральной специализации микротрубочек в общем процессе «распаковки» Информации из эфирных конструкций ОДС и ФЛК нейроны головного мозга функционально подразделяются на афферентные (чувствительные, воспринимающие Информацию), эфферентные (передающие Информацию от нервного центра к исполнительным органам или другим центрам нервной системы) и вставочные (интернейроны, осуществляющие коммуникацию внутри каждого отдела мозга: обрабатывают Информацию, получаемую от афферентных нейронов, и передают её другим вставочным или эфферентным нейронам). Все они – вибрационно и сложнофункционально – очень тесно связаны с Формо-Творцами первых трёх пар ИИССИИДИ-Центров.
5.147. Внутри каждая микротрубочка полая (как цилиндр), так как все глобулы концентрически расположены («упакованы») по её диаметру, который не меняется (24-25 нм по наружному и 14-15 нм по внутреннему контуру). При рассмотрении функций нейрофибрилл очень важным обстоятельством является то, что протеин тубулин представляет собой димер, состоящий из α- и ß-тубулина, благодаря чему на одном конце микротрубочки осуществляется самосборка (пополнение новыми фрагментами), в то время как на другом конце происходит её разборка (транспортировка фрагмента в другую нейрофибриллу). Примечательность свойств этих поляризованных молекул протеинов-димеров заключается ещё и в том, что в определённых участках формируемой ими структуры электроны проявляют все признаки квантовой запутанности (возникновение и исчезновение квантовой когерентности), в результате чего характер поведения одного из электронов существенно влияет на состояние другого электрона (даже в случае их пространственного дистанцирования).
5.148. Обладая таким универсальным свойством, электроны всех молекул белков тубулинов и ассоциированных с ними МАР-белков, последовательно синтезируемых в нейрофибриллах, волновым путём симультанно разносят (транслируют) сразу по всем микротрубочкам остальных клеток организма свойственную им фрагментарную Энерго-Информацию, которую другие био-Творцы либо резонационно используют для характерной им биохимической реализации, либо просто игнорируют, в результате чего протеины в микротрубочках быстро распадаются. Учитывая, что каждый из 86 млрд. нейронов спонтанно генерирует в нейронную сеть мозга свои собственные импульсы, ежесекундно по всем нейронным сетям хаотически распространяются миллиарды всевозможных шумовых, фоновых информационных импульсов, лишь мизерная часть которых подвергается реальной обработке конкретными био-Творцами.
5.149. Димерные свойства белков-тубулинов являются главной причиной динамической нестабильности нейрофибрилл, которая выражается в свойственных им процессах непрерывной полимеризации и деполимеризации, что не позволяет им находиться в каком-то одном устойчивом состоянии: рост микротрубочки может лишь относительно стабилизироваться, когда процессы деполимеризации будут визуально уравновешиваться процессами полимеризации (то есть когда объём ретранслированных фрагментов Энерго-Информации будет восполняться из ОДС фрагментами новой Информации). Через эту важную особенность отражается одно из основных свойств любой Фокусной Динамики – непрерывная мультиполяризационная смена одних квантовых «сдвигов» на качественно другие, что составляет собой основу осуществления Формо-Творцами всех их субъективных амицирационных (перефокусировочных) процессов.
5.150. Нейроны могут функционировать в двух режимах: либо самопроизвольно генерировать импульсы, либо обрабатывать сигналы, поступающие от других нейронов, чтобы передать результирующий импульс дальше (при этом лишь около 20% синапсов в коре являются тормозящими, в то время как остальные – стимулирующими). На эти процессы расходуется собственная, но предварительно накопленная другими внутриклеточными ФС (например, лизосомами, митохондриями), декогерентная Энергия каждого нейрона, которая – непрерывно локально высвобождаясь – употребляется для запуска аналогичных процессов в соседних клетках.
5.151. Таким образом, всю нейронную сеть мозга каждого индивидуума, структурированную примерно 86 миллиардами самых разнообразных по строению и функциям нейронов (грушевидных, звездчатых, веретенообразных, шаровидных, пирамидных и так далее), вполне справедливо можно отнести к самоорганизующимся активным средам, в различных условиях эксгиберации которых (за счёт симультанной мультиполяризации Фокусных Динамик всех био-Творцов) может самопроизвольно зарождаться, развиваться и реализовываться трансфинитное множество принципиально новых свойств, качеств и творческих возможностей. На этой эгллеролифтивной энергоинформационной основе в различных режимах Пространства-Времени («сценариях» развития) симультанно сформировалось и проявилось всё бесчисленное многообразие НУУ-ВВУ-Конфигураций всевозможных «личностных» Интерпретаций.
5.152. Био-Творцы микротрубочек разных нейронов могут симультанно – как волновым, так и химическим образом – быть взаимосвязаны между собой и целенаправленно корректировать происходящие в них (и с ними самими) процессы, которые становятся возможными лишь при наличии био-Творцов молекул гуанозинтрифосфата (связанных с ß-тубулином), представляющих не только в цитоструктуре каждого нейрона, но также и в структуре любой иной эукариотической клетки реализационные Интересы ядерных Формо-Творцов генома. Надо отметить, что гидролизные функции био-Творцов ГТФ – в бирвуляртном отношении – являются более специфичными, чем функции био-Творцов АТФ, чьи Фокусные Динамики отражают полипротоформные Интересы представителей множества типов бирвуляртности, структурирующих наш биологический организм.
5.153. Эти био-Творцы (ГТФ) постоянно направляют и соответствующим образом корректируют все процессы сборки и разборки нейрофибрилл, обеспечивая их необходимым для этого Энерго-Потенциалом. Причём, в зависимости от конкретной ситуации и предоставляемых ею реализационных возможностей, декогерентная Энергия химических реакций гуанозинтрифосфата может быть направлена не только на решение каких-то Интересов, свойственных непосредственно Творцам-регуляторам ллууввумической Схемы Синтеза (накопление Опыта человеческих переживаний), но также и на прочие разнопротоформные реализации ФД био-Творцов всего множества ансамблей клеток, которые обеспечивают целостное и согласованное функционирование всех участников нашего белково-нуклеинового комплекса.
5.154. То есть для получения и последующей передачи Энерго-Информации, поступающей из ОДС или ФЛК и симультанно транспортируемой во все системы нашего организма через непрерывный процесс деформации (поляризации и деполяризации) нейронных микротрубочек (разнопротоформные био-Творцы которых химически декодируют поступающие к ним информационные сигналы, переводя их в электромагнитные импульсы), любой Форме Самосознания, состоящей из клеток, совсем не обязательно иметь собственную разветвлённую нервную систему (по типу периферической нервной системы человека), так как ретрансляция между микротрубочками абсолютно всех клеток симультанно осуществляется нутационным (волновым) путём. Эффективность конкретной реакции разнопротоформных био-Творцов каждой клетки организма на полученный таким путём импульс (а значит, и степень их влияния на общее качественное содержимое ФД «личности») полностью зависит от наличия в ней таких важных энергетических носителей, как АТФ и ГТФ, распределением которых в клетке занимаются геномные Формо-Творцы.
5.155. Поскольку содержимое (в виде СФУУРММ-Формы) каждого информационного посыла, попадая в субтеррансивные условия цитосистемы клетки, подвергается множественной разнокачественной интерпретации через мультиполяризационные свойства Фокусных Динамик различных протоформных био-Творцов, обрабатывающих эту Информацию и реализационно мотивированных общим набором «текущих» (от самых деплиативных до самых амплиативных) Интересов Творцов-интерпретаторов ДНК, то и Фокусная Динамика самой «личности» при реализации будет обладать теми же мультиполяризационными свойствами: каждому варианту субтеррансивной «распаковки» ФД будет соответствовать свой собственный «сценарий» развития. Амицирация в тот или иной «сценарий» зависит от качественности ф-Конфигурации фокусируемого вами НУУ-ВВУ-Формо-Типа, то есть от того, какие из био-Творцов конкретно задействованы в цепочке приёма Информации и ретрансляции её био-Творцам всех остальных микротрубочек в виде уже специфически маркированной протеином Энерго-Информации.
5.156. В теле каждого нейрона основная масса нейрофибрилл ориентирована радиально от центра своего образования к периферии клетки, в то время как другая их часть ориентирована в противоположном направлении. В месте разграничения тела нейрона и его отростка (вдоль аксона) образуется некая компактная структура, функционально ориентированная на конец отростка и состоящая из 15-30 стабильных микротрубочек специфического качества. Эта конструкция представляет собой своеобразную распределительную станцию, через которую частично уже отсортированная Энерго-Информация транслируется из тел нейронов разных клеток далее – к нейронам соответствующих участков коры и подкорки головного мозга (в ФД их био-Творцов), а затем – по всей нервной системе.
5.157. Сами же отростки нейрона вдоль всей длины своей оси упорядоченно структурированы несколькими разновидностями микротрубочек, большинство которых располагается поодиночке, постоянно «урезающимся» концом в сторону синапса (аксонного окончания), где прямые микротрубочки, «освободившись» от большей части своего содержимого в момент постсинаптической трансляции информационного сигнала, в различной степени деформируются (некоторые же, не будучи использованы по назначению – в силу некорректного декодирования или отсутствия ВВУ-Фактора, – изгибаются в аксонном окончании и транспортируют своё органическое содержимое обратно в тело нейрона для последующей химической переработки и использования в новой нейрофибрилле).
5.158. Микротрубочки могут быть как длинными (кажущимися стабильными и неподвижными), так и короткими (мобильными), а, в зависимости от типа нервной клетки и своего расположения в нейроне и его отростках, их длина может составлять от нескольких десятков нанометров до десятков микрон, что напрямую связано с особенностями их функциональности: длинные нейрофибриллы являются промежуточными накопителями Энерго-Информации, поступающей в систему Восприятия, а короткие – активными посредниками и переносчиками отдельных её фрагментов (УУ-ВВУ-копий), из которых затем Фокусными Динамиками различных био-Творцов головного мозга в информационном «пространстве» «личностного» Самосознания организуются абсолютно все СФУУРММ-Формы. В среднем каждая трубочка выполняет в нейроне свойственную ей функцию в течение временного промежутка от нескольких минут до получаса, качественно и структурно изменяясь при этом по мере происходящих в ней энергоинформационных трансформаций. Основную роль в резонационной организации этого процесса играют Формы Самосознаний различных нейромедиаторов, а также био-Творцов множества качественно специализированных (по реализационному характеру всевозможных вибрационных Уровней) низко- и высокомолекулярных белков.
5.159. Качественная трансформация микротрубочек из одного вида в другой осуществляется благодаря наличию в цитоплазме ФС специфических ферментов – катанина и спастина: первый дифференцирует длинные микротрубочки на короткие подвижные фрагменты (длиной около 10 нм), которые, выполняя характерные им функции по передаче определённой Энерго-Информации, перемещаются внутри цитоплазмы и по отросткам нейрона на расстояния в десятки и сотни микрон, в то время как второй присоединяет «отрезанные» фрагменты к нужному (приспособленному для непрерывного наращивания) концу другой длинной нейрофибриллы, от противоположного (постоянно убывающего) конца которой катанин тут же «отрезает» соответствующий фрагмент, чтобы, транспортировав его на расстояние 20-60 нм, немедленно присоединить к наращиваемому концу другой микротрубочки.
5.160. То есть в нейроне и его отростках все микротрубочки постоянно подвержены процессу дифференциации (деполимеризации – избавлению от наиболее диссонационных для данной СФУУРММ-Формы УУ-ВВУ-фрагментов), качественного обновления (полимеризации – пополнению более коварллертными УУ-ВВУ-фрагментами) и последующей резонационной трансформации по цитоплазме нейрона к местам своего непосредственного биохимического декодирования био-Творцами коры и подкорки: с одного конца каждая длинная трубочка постоянно урезается на определённый УУ-ВВУ-фрагмент, в то время как другой её конец непрерывно пополняется более коварллертными УУ-ВВУ-фрагментами всё новой и новой Энерго-Информации. За счёт симультанности этого процесса и создаётся эффект относительного визуального постоянства размера длинных микротрубочек.
5.161. Следует отметить, что нейрофибриллы с похожим по качественности информационным содержанием, отражающим реализационные особенности СФУУРММ-Форм какого-то определённого каузального «Канала» (или коварллертных подуровней конкретной пары ИИССИИДИ-Центров), «последовательно» формируются в нейрофиламенты – длинные (от 50-60 нм до одного и более микрон) и субтеррансивные по своим электромагнитным параметрам цепочки нейрофиламентозных белков (основной из которых – актин – по своим сократительным свойствам близок тубулину). Надо понимать, что последовательность, о которой идёт речь, нельзя отнести к прямой, потому что нейрофибриллы не стыкуются в нейрофиламенты, как вагоны в железнодорожный состав, ведь «вагоны» тоже могут быть разными в зависимости от «перевозимой» ими Информации, и формирование нейрофиламентов происходит на базе частотного совпадения более коварллертной Информации: нейрофибриллы с Информацией основного типа (наиболее близкие по своему «содержанию») «превращаются» в нейрофиламенты.
5.162. Этот процесс пока не может быть доступен исследованиям учёных, так как то, что мы визуально воспринимаем как «материю», представляет собой «проекции» более высоких энергий в условиях более высокой плотности (не вибрационной, а материальных – «физических» – взаимосвязей). И поскольку нейрофиламенты представляют собой микромиры (то есть совсем другие условия ПВК, не те, которые мы с вами можем наблюдать и которые можем реализовывать в нашей ФД), то они являются посредниками между более амплиативными ПВК (спектр проявления которых включает высшие Уровни 3-4-мерного и низшие Уровни 4-5-мерного диапазонов мерностей) и «нашими» – ирккуллигренными – Реальностями. Достаточно «присмотреться» (в глубинной Медитации, например) к микромиру и «спроецировать» свойственные ему взаимосвязи на более «глобальные» структуры макромира, чтобы убедиться в том, что микротрубочки не переходят в нейрофиламенты автоматически, а происходит их реорганизация, необходимая для достижения совместимости структурирующей их разнокачественной Информации. Эта Информация перераспределяется по частотным и качественным параметрам и уже в этих покачественных преобразованиях формируются нейрофиламенты: когда собирается более-менее резонационно идентичная Информация, принадлежащая одним уровням (одному смысловому ряду), и организуется через потенциал нейрофиламентов, то на выходе получается общая СФУУРММ-Форма – готовый продукт Творцов данного нейрона для передачи следующему нейрону. При этом нейрофиламенты, хотя и кажутся визуально более стабильными, чем микротрубочки, также, по мере использования био-Творцами свойственной им Энерго-Информации, подвержены постоянной корректировке и обновлению с помощью Форм Самосознаний специальных ферментов.
5.163. Именно нейрофиламенты представляют собой ту первичную материальную протооснову Энерго-Информации, которая (после биохимического декодирования её био-Творцами коры, подкорки и других систем организма) конкатенационно формируется в структуре генома и психоментально проявляется в ФД Самосознания «личности» как её некое субъективное Представление о чём-то или о ком-то. Точно так же, как и формируемые на их основе СФУУРММ-Формы, которые невозможно резко дифференцировать и отделить друг от друга, нейрофиламенты глубоко связаны и между собой, и с отдельными микротрубочками, образуя (вместе с остальными компонентами клетки) в теле и отростках нейронов очень сложную энергоинформационную ретрансляционную сеть, одновременно выполняющую функции материальной (цитоскелетной) основы как каждого нейрона, так и любой другой клетки нашего организма.
5.164. По мере реализации в ФД «личности» содержимого определённых микротрубочек и нейрофиламентов (через соответствующие психоментальные переживания, физические ощущения и действия), в её «субтеррансивной ОДС» происходит последовательный Синтез определённого Опыта, который фиксируется и соответствующим образом маркируется (клексуется и помечается как уже отработанный) Формо-Творцами гиппокампа, а впоследствии интерпретируется нами как один из элементов нашей «долгосрочной памяти». При болезни Альцгеймера, рассеянном склерозе и других нарушениях в центральной нервной системе реализационные приоритеты геномных Формо-Творцов качественно меняются, в результате чего функциональные возможности био-Творцов головного мозга также существенно понижаются, поскольку, в силу дефицита ФС гуанозинтрифосфата (главного для нейронов источника декогерентной Энергии), они уже не способны сразу (или достаточно быстро) резонационно «распаковывать» (биохимически обрабатывать) большое количество непрерывно поступающих к ним СФУУРММ-Форм.
5.165. Существенная часть этой Информации, будучи уже воспринятой тем или иным образом (зрительным, слуховым, обонятельным, вкусовым, тактильным), остаётся некоторое время в микротрубочках в неопределённом состоянии, то есть пребывает без дальнейших реализационных последствий, что при данном заболевании выражается в первоочередном поражении трансэнторинальной коры головного мозга (связующее звено между ассоциативными областями неокортекса), а позднее – в вовлечении в патологический процесс био-Творцов гиппокампа и далее – коры больших полушарий. Поэтому в телах и отростках нейронов образуются своеобразные информационные «пробки», «завалы», «заторы» (так называемые «тельца Хирано» – избыточно сконцентрированные актиновые микрофиламенты в проксимальных дендритах), состоящие из «нераспакованных» должным образом и своевременно необработанных био-Творцами нейрофиламентов.
5.166. Сами же микротрубочки в таких ситуациях (появления и накапливания нейрофибриллярных клубков, образованных избыточно фосфорилированными тау-белками) достаточно сильно дестабилизируются, что неизбежно влечёт за собой искажение передаваемой ими Энерго-Информации. Множество необработанных нейрофиламентов, так и не дождавшись освобождения от структурирующих их нейрофиламентозных белков, как бы «растворяются» (то есть «перепроецируются» в более благоприятные условия других «сценариев») или качественно переориентируются (переформатируются другим Смыслом, более доступным при имеющемся ГТФ-дефиците), в результате чего концентрация микротрубочек вдоль длинной оси отростков нейрона уменьшается.
5.167. Одной из главных причин необходимости в нашей жизни сна и хотя бы короткого переключения внимания при выполнении нами сложной умственной работы на более простой или отвлечённый вид деятельности является именно предоставление био-Творцам микротрубочек и структурируемых ими нейрофиламентов реальных возможностей для своевременной обработки и транспортировки биохимически маркированной ими Энерго-Информации в соответствующие её реализационному назначению участки белково-нуклеинового комплекса. В противном случае в клетках происходит постепенное накопление сверхфосфолирированных тау-белков, что чревато возникновением в организме различных дисфункциональных состояний (например, предстарческой деменции, синдромов хронической утомляемости, Альцгеймера и других).




